

Structure of a baculovirus sulfhydryl oxidase, a highly divergent member of the erv flavoenzyme family
- PMID: 21752922
- PMCID: PMC3165737
- DOI: 10.1128/JVI.05149-11
Structure of a baculovirus sulfhydryl oxidase, a highly divergent member of the erv flavoenzyme family
Abstract
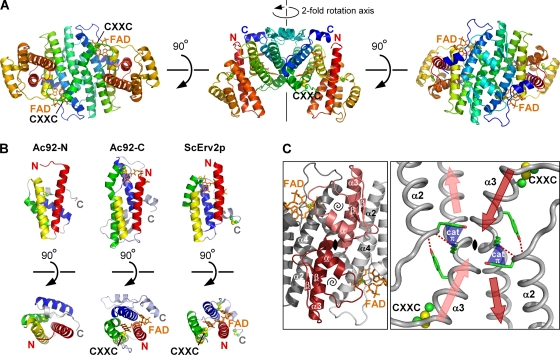
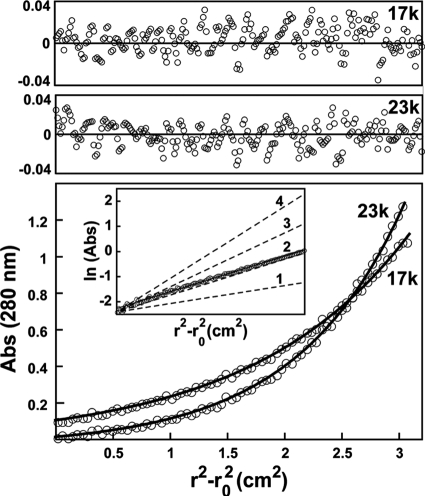
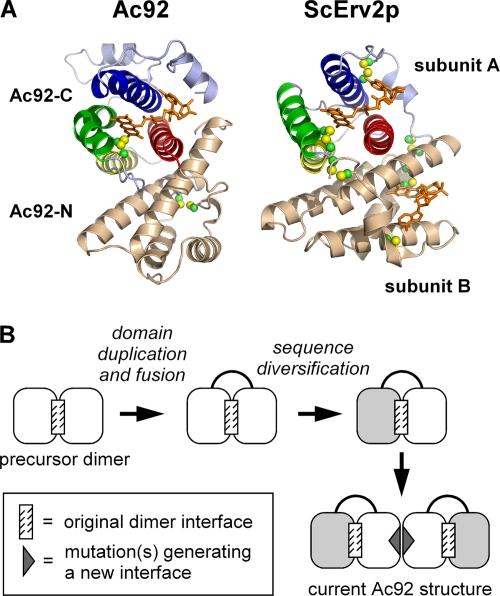
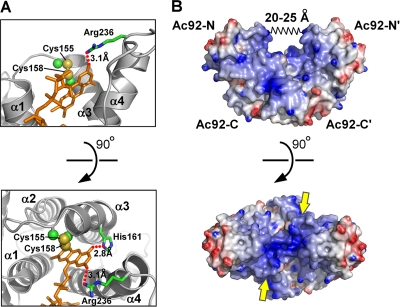
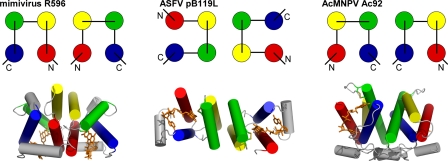
Genomes of nucleocytoplasmic large DNA viruses (NCLDVs) encode enzymes that catalyze the formation of disulfide bonds between cysteine amino acid residues in proteins, a function essential for the proper assembly and propagation of NCLDV virions. Recently, a catalyst of disulfide formation was identified in baculoviruses, a group of large double-stranded DNA viruses considered phylogenetically distinct from NCLDVs. The NCLDV and baculovirus disulfide catalysts are flavin adenine dinucleotide (FAD)-binding sulfhydryl oxidases related to the cellular Erv enzyme family, but the baculovirus enzyme, the product of the Ac92 gene in Autographa californica multiple nucleopolyhedrovirus (AcMNPV), is highly divergent at the amino acid sequence level. The crystal structure of the Ac92 protein presented here shows a configuration of the active-site cysteine residues and bound cofactor similar to that observed in other Erv sulfhydryl oxidases. However, Ac92 has a complex quaternary structural arrangement not previously seen in cellular or viral enzymes of this family. This novel assembly comprises a dimer of pseudodimers with a striking 40-degree kink in the interface helix between subunits. The diversification of the Erv sulfhydryl oxidase enzymes in large double-stranded DNA viruses exemplifies the extreme degree to which these viruses can push the boundaries of protein family folds.
Figures
References
-
- Collaborative Computational Project, Number 4 1994. The CCP4 suite: programs for protein crystallography. Acta Crystallogr. D Biol. Crystallogr. 50:760–763 - PubMed
-
- Allen S., Balabanidou V., Sideris D. P., Lisowsky T., Tokatlidis K. 2005. Erv1 mediates the Mia40-dependent protein import pathway and provides a functional link to the respiratory chain by shuttling electrons to cytochrome c. J. Mol. Biol. 353:937–944 - PubMed
-
- Braunagel S. C., Summers M. D. 2007. Molecular biology of the baculovirus occlusion-derived virus envelope. Curr. Drug Targets 8:1084–1095 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
