

The same binding in contour integration and crowding
- PMID: 21757504
- PMCID: PMC3624759
- DOI: 10.1167/11.8.10
The same binding in contour integration and crowding
Abstract
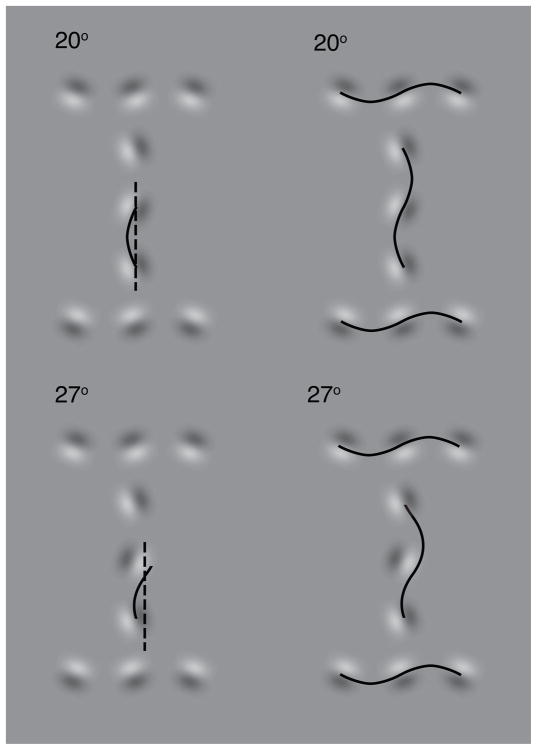
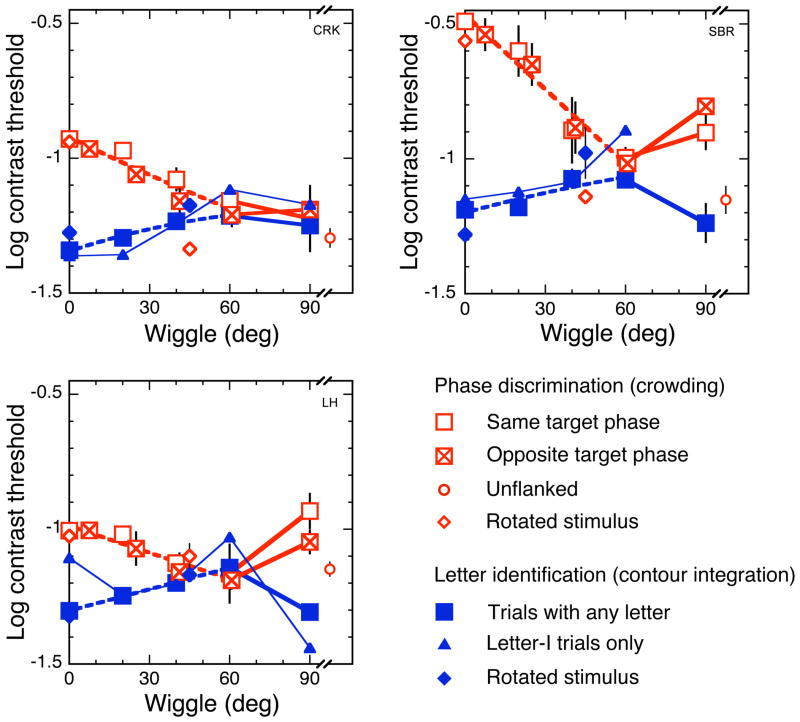
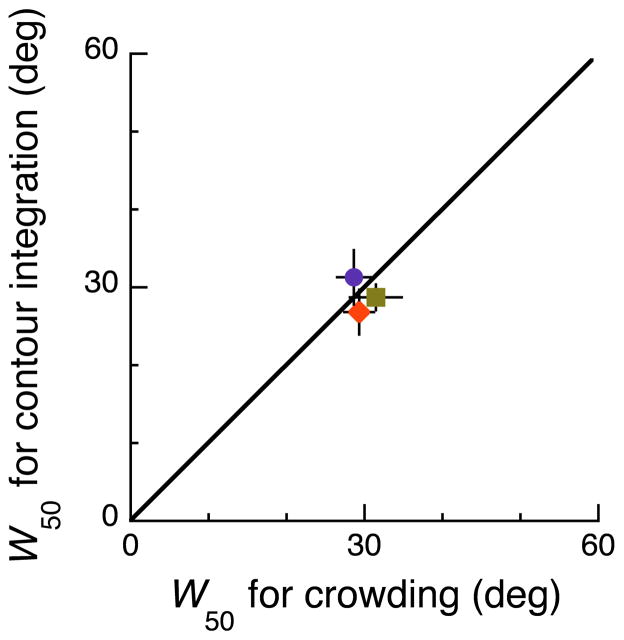
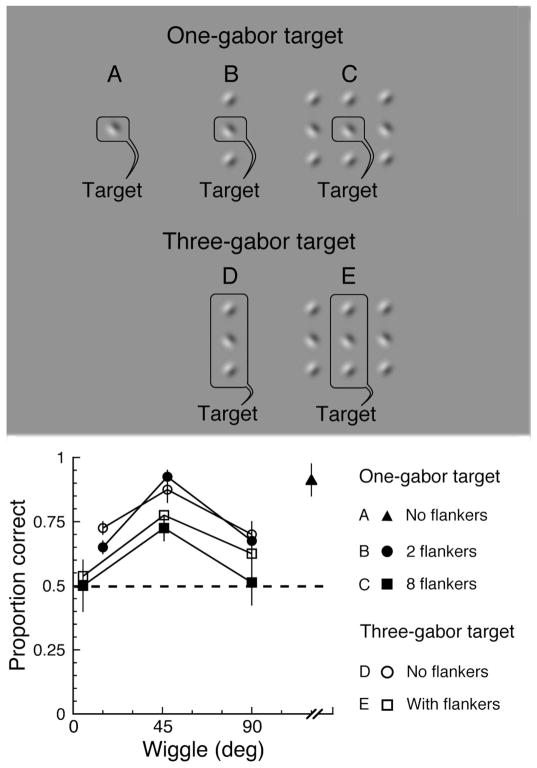
Binding of features helps object recognition in contour integration but hinders it in crowding. In contour integration, aligned adjacent objects group together to form a path. In crowding, flanking objects make the target unidentifiable. However, to date, the two tasks have only been studied separately. K. A. May and R. F. Hess (2007) suggested that the same binding mediates both tasks. To test this idea, we ask observers to perform two different tasks with the same stimulus. We present oriented grating patches that form a "snake letter" in the periphery. Observers report either the identity of the whole letter (contour integration task) or the phase of one of the grating patches (crowding task). We manipulate the strength of binding between gratings by varying the alignment between them, i.e., the Gestalt goodness of continuation, measured as "wiggle." We find that better alignment strengthens binding, which improves contour integration and worsens crowding. Observers show equal sensitivity to alignment in these two very different tasks, suggesting that the same binding mechanism underlies both phenomena. It has been claimed that grouping among flankers reduces their crowding of the target. Instead, we find that these published cases of weak crowding are due to weak binding resulting from target-flanker misalignment. We conclude that crowding is mediated solely by the grouping of flankers with the target and is independent of grouping among flankers.
Figures

Similar articles
-
The role of crowding in contextual influences on contour integration.J Vis. 2012 Jul 9;12(7):3. doi: 10.1167/12.7.3. J Vis. 2012. PMID: 22776847
-
Object crowding.J Vis. 2011 May 25;11(6):10.1167/11.6.19 19. doi: 10.1167/11.6.19. J Vis. 2011. PMID: 21613388 Free PMC article.
-
Illusory contour formation survives crowding.J Vis. 2012 Jun 12;12(6):15. doi: 10.1167/12.6.15. J Vis. 2012. PMID: 22693333
-
Ladder contours are undetectable in the periphery: a crowding effect?J Vis. 2007 Oct 29;7(13):9.1-15. doi: 10.1167/7.13.9. J Vis. 2007. PMID: 17997637
-
The uncrowded window of object recognition.Nat Neurosci. 2008 Oct;11(10):1129-35. doi: 10.1038/nn.2187. Nat Neurosci. 2008. PMID: 18828191 Free PMC article. Review.
Cited by
-
Emergence of crowding: The role of contrast and orientation salience.J Vis. 2021 Oct 5;21(11):20. doi: 10.1167/jov.21.11.20. J Vis. 2021. PMID: 34709355 Free PMC article.
-
Similar Sensitivity to Ladder Contours in Macular Degeneration Patients and Controls.PLoS One. 2015 Jul 14;10(7):e0128119. doi: 10.1371/journal.pone.0128119. eCollection 2015. PLoS One. 2015. PMID: 26171784 Free PMC article.
-
Seven Myths on Crowding and Peripheral Vision.Iperception. 2020 May 19;11(3):2041669520913052. doi: 10.1177/2041669520913052. eCollection 2020 May-Jun. Iperception. 2020. PMID: 32489576 Free PMC article. Review.
-
Combining S-cone and luminance signals adversely affects discrimination of objects within backgrounds.Sci Rep. 2016 Feb 9;6:20504. doi: 10.1038/srep20504. Sci Rep. 2016. PMID: 26856308 Free PMC article. Clinical Trial.
-
Visual crowding cannot be wholly explained by feature pooling.J Exp Psychol Hum Percept Perform. 2014 Jun;40(3):1022-33. doi: 10.1037/a0035377. Epub 2013 Dec 23. J Exp Psychol Hum Percept Perform. 2014. PMID: 24364703 Free PMC article.
References
-
- Andriessen JJ, Bouma H. Eccentric vision: adverse interactions between line segments. Vision Research. 1976;16 (1):71– 78. - PubMed
-
- Appelle S. Perception and discrimination as a function of stimulus orientation: the “oblique effect” in man and animals. Psychological Bulletin. 1972;78 (4):266– 278. - PubMed
-
- Bouma H. Interaction effects in parafoveal letter recognition. Nature. 1970;226:177– 178. - PubMed
-
- Brainard DH. The Psychophysics Toolbox. Spatial Vision. 1997;10(4):433– 436. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
