

Neuroprotective role of PrPC against kainate-induced epileptic seizures and cell death depends on the modulation of JNK3 activation by GluR6/7-PSD-95 binding
- PMID: 21757544
- PMCID: PMC3164453
- DOI: 10.1091/mbc.E11-04-0321
Neuroprotective role of PrPC against kainate-induced epileptic seizures and cell death depends on the modulation of JNK3 activation by GluR6/7-PSD-95 binding
Abstract
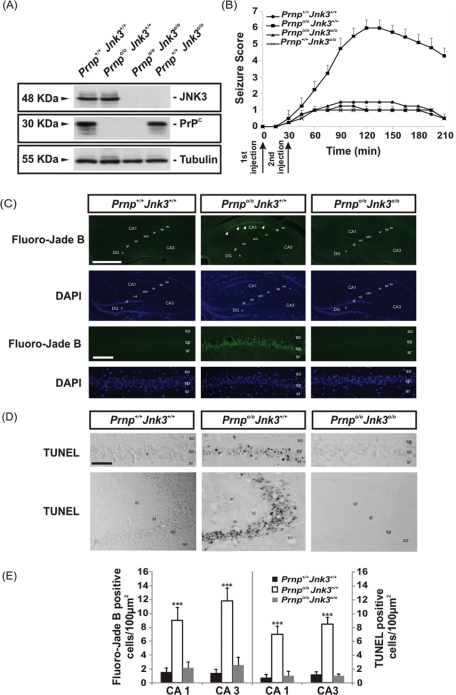
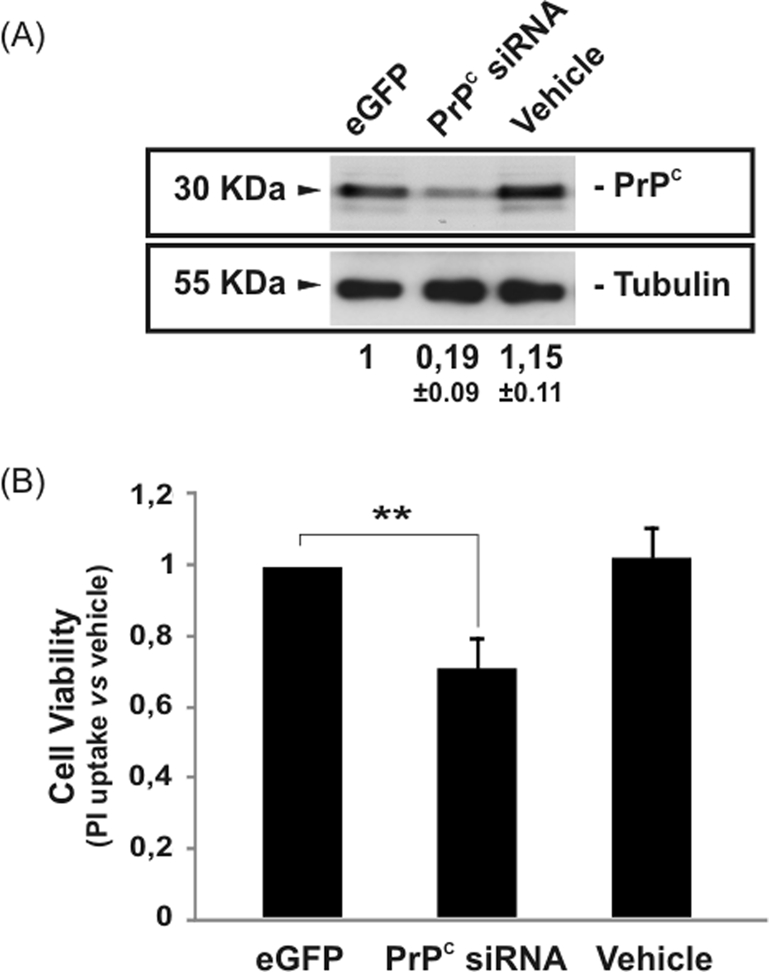
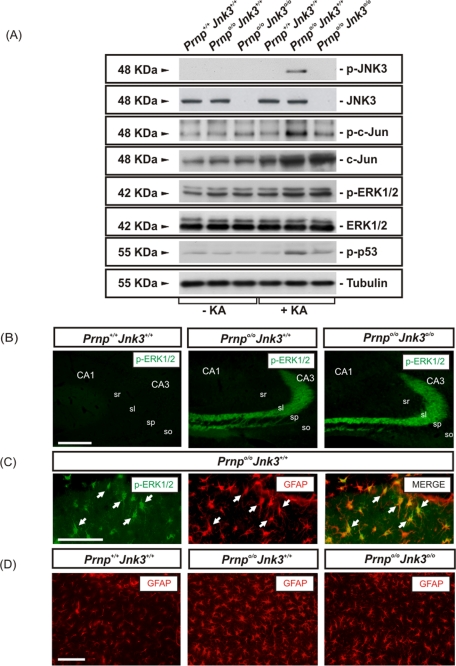
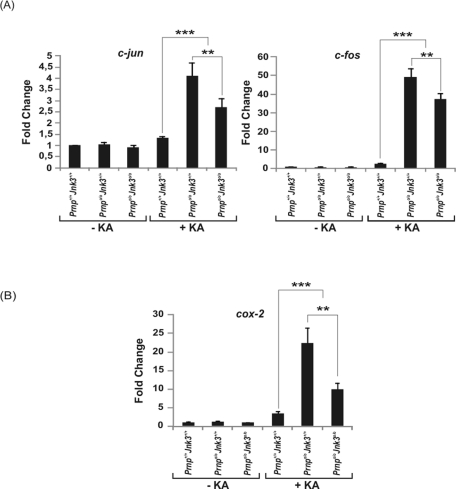
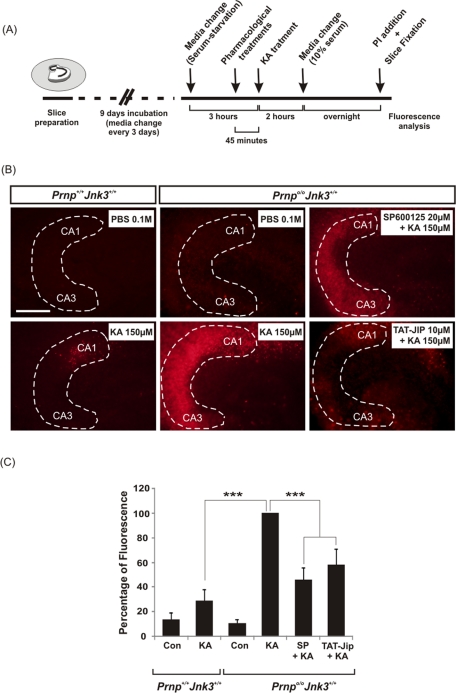
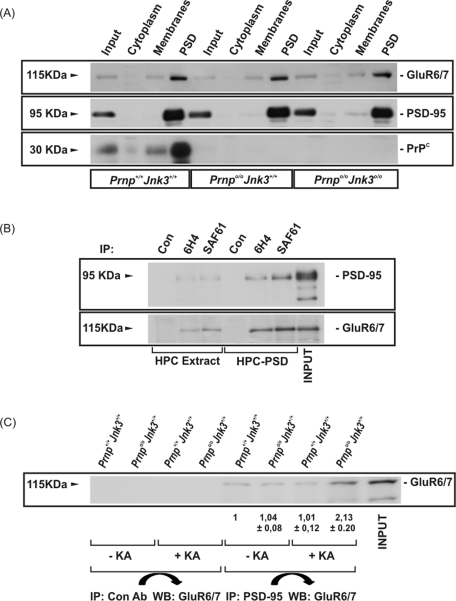
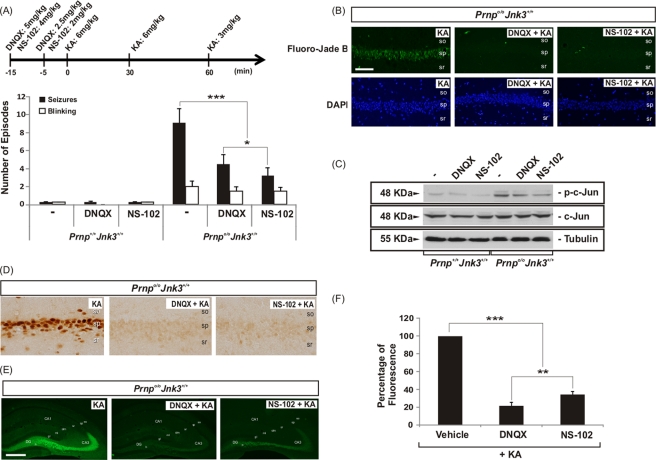
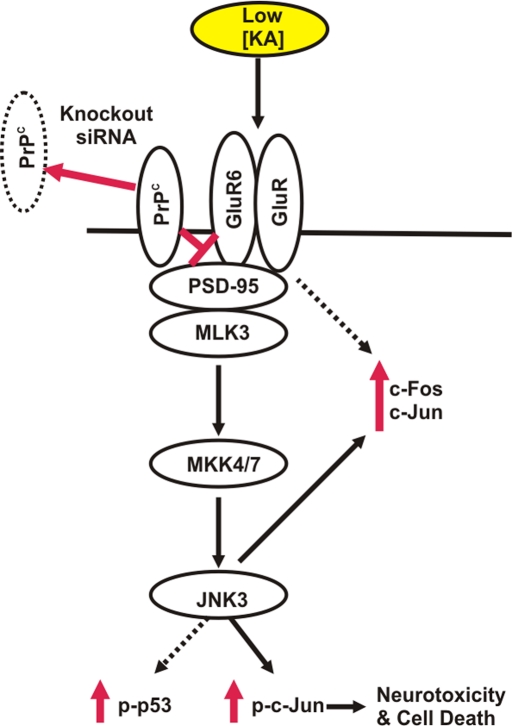
Cellular prion protein (PrP(C)) is a glycosyl-phosphatidylinositol-anchored glycoprotein. When mutated or misfolded, the pathogenic form (PrP(SC)) induces transmissible spongiform encephalopathies. In contrast, PrP(C) has a number of physiological functions in several neural processes. Several lines of evidence implicate PrP(C) in synaptic transmission and neuroprotection since its absence results in an increase in neuronal excitability and enhanced excitotoxicity in vitro and in vivo. Furthermore, PrP(C) has been implicated in the inhibition of N-methyl-d-aspartic acid (NMDA)-mediated neurotransmission, and prion protein gene (Prnp) knockout mice show enhanced neuronal death in response to NMDA and kainate (KA). In this study, we demonstrate that neurotoxicity induced by KA in Prnp knockout mice depends on the c-Jun N-terminal kinase 3 (JNK3) pathway since Prnp(o/o)Jnk3(o/o) mice were not affected by KA. Pharmacological blockage of JNK3 activity impaired PrP(C)-dependent neurotoxicity. Furthermore, our results indicate that JNK3 activation depends on the interaction of PrP(C) with postsynaptic density 95 protein (PSD-95) and glutamate receptor 6/7 (GluR6/7). Indeed, GluR6-PSD-95 interaction after KA injections was favored by the absence of PrP(C). Finally, neurotoxicity in Prnp knockout mice was reversed by an AMPA/KA inhibitor (6,7-dinitroquinoxaline-2,3-dione) and the GluR6 antagonist NS-102. We conclude that the protection afforded by PrP(C) against KA is due to its ability to modulate GluR6/7-mediated neurotransmission and hence JNK3 activation.
Figures
References
-
- Aguzzi A, Calella AM. Prions: protein aggregation and infectious diseases. Physiol Rev. 2009;89:1105–1152. - PubMed
-
- Aguzzi A, Heikenwalder M, Polymenidou M. Insights into prion strains and neurotoxicity. Nat Rev Mol Cell Biol. 2007;8:552–561. - PubMed
-
- Alford S, Grillner S. CNQX and DNQX block non-NMDA synaptic transmission but not NMDA-evoked locomotion in lamprey spinal cord. Brain Res. 1990;506:297–302. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
