

Phylogeny-wide analysis of social amoeba genomes highlights ancient origins for complex intercellular communication
- PMID: 21757610
- PMCID: PMC3205573
- DOI: 10.1101/gr.121137.111
Phylogeny-wide analysis of social amoeba genomes highlights ancient origins for complex intercellular communication
Abstract
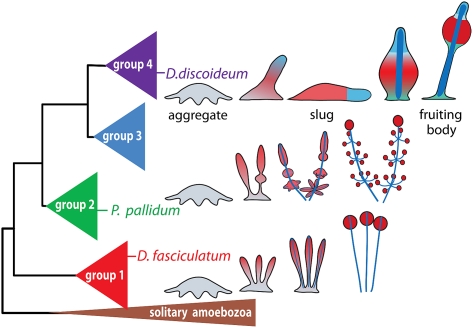
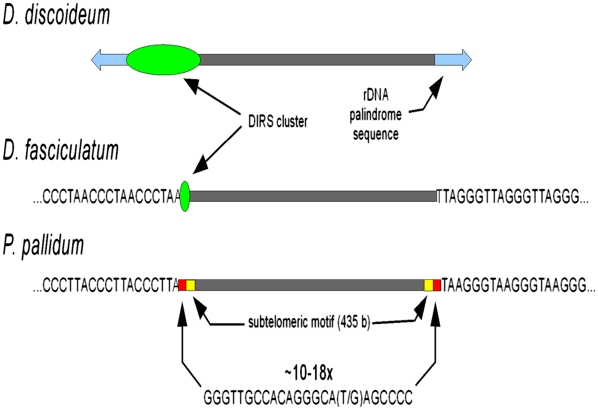
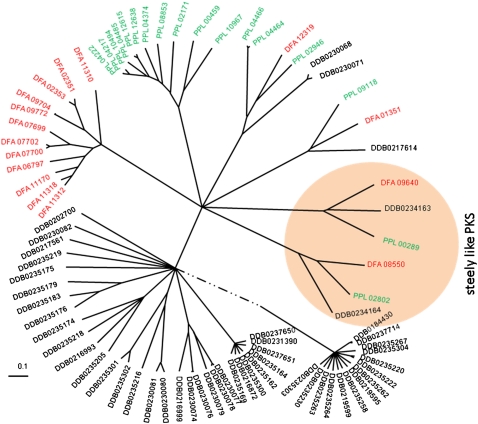
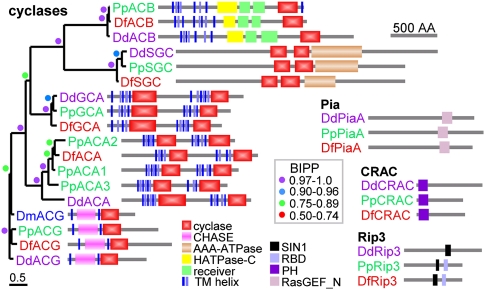
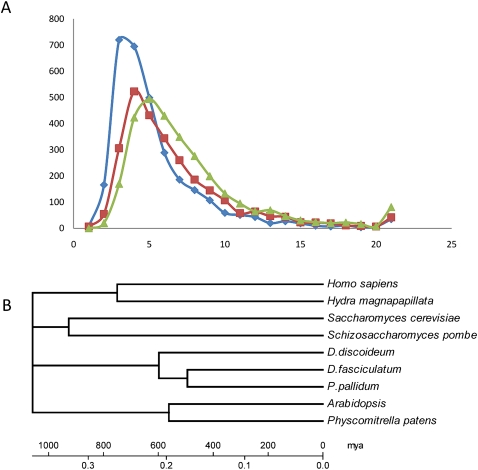
Dictyostelium discoideum (DD), an extensively studied model organism for cell and developmental biology, belongs to the most derived group 4 of social amoebas, a clade of altruistic multicellular organisms. To understand genome evolution over long time periods and the genetic basis of social evolution, we sequenced the genomes of Dictyostelium fasciculatum (DF) and Polysphondylium pallidum (PP), which represent the early diverging groups 1 and 2, respectively. In contrast to DD, PP and DF have conventional telomere organization and strongly reduced numbers of transposable elements. The number of protein-coding genes is similar between species, but only half of them comprise an identifiable set of orthologous genes. In general, genes involved in primary metabolism, cytoskeletal functions and signal transduction are conserved, while genes involved in secondary metabolism, export, and signal perception underwent large differential gene family expansions. This most likely signifies involvement of the conserved set in core cell and developmental mechanisms, and of the diverged set in niche- and species-specific adaptations for defense and food, mate, and kin selection. Phylogenetic dating using a concatenated data set and extensive loss of synteny indicate that DF, PP, and DD split from their last common ancestor at least 0.6 billion years ago.
Figures
References
-
- Anjard C, Loomis WF 2006. GABA induces terminal differentiation of Dictyostelium through a GABAB receptor. Development 133: 2253–2261 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous