

Characterization of the deoxynucleotide triphosphate triphosphohydrolase (dNTPase) activity of the EF1143 protein from Enterococcus faecalis and crystal structure of the activator-substrate complex
- PMID: 21757692
- PMCID: PMC3190883
- DOI: 10.1074/jbc.M111.250456
Characterization of the deoxynucleotide triphosphate triphosphohydrolase (dNTPase) activity of the EF1143 protein from Enterococcus faecalis and crystal structure of the activator-substrate complex
Abstract
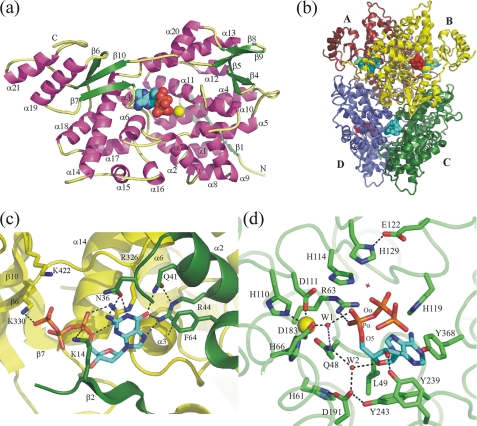
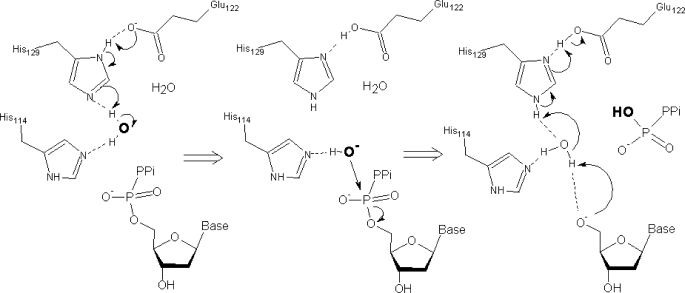
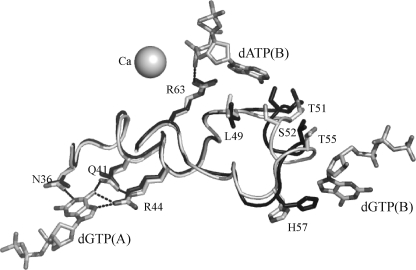
The EF1143 protein from Enterococcus faecalis is a distant homolog of deoxynucleotide triphosphate triphosphohydrolases (dNTPases) from Escherichia coli and Thermus thermophilus. These dNTPases are important components in the regulation of the dNTP pool in bacteria. Biochemical assays of the EF1143 dNTPase activity demonstrated nonspecific hydrolysis of all canonical dNTPs in the presence of Mn(2+). In contrast, with Mg(2+) hydrolysis required the presence of dGTP as an effector, activating the degradation of dATP and dCTP with dGTP also being consumed in the reaction with dATP. The crystal structure of EF1143 and dynamic light scattering measurements in solution revealed a tetrameric oligomer as the most probable biologically active unit. The tetramer contains four dGTP specific allosteric regulatory sites and four active sites. Examination of the active site with the dATP substrate suggests an in-line nucleophilic attack on the α-phosphate center as a possible mechanism of the hydrolysis and two highly conserved residues, His-129 and Glu-122, as an acid-base catalytic dyad. Structural differences between EF1143 apo and holo forms revealed mobility of the α3 helix that can regulate the size of the active site binding pocket and could be stabilized in the open conformation upon formation of the tetramer and dGTP effector binding.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
