

Agonist-directed interactions with specific beta-arrestins determine mu-opioid receptor trafficking, ubiquitination, and dephosphorylation
- PMID: 21757712
- PMCID: PMC3173119
- DOI: 10.1074/jbc.M111.248310
Agonist-directed interactions with specific beta-arrestins determine mu-opioid receptor trafficking, ubiquitination, and dephosphorylation
Abstract
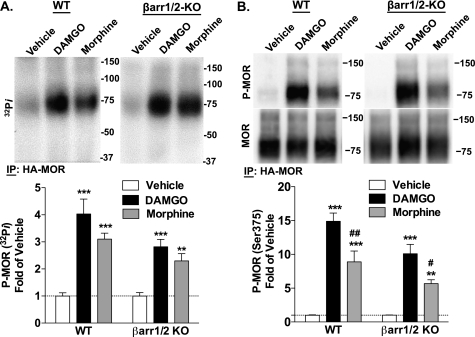
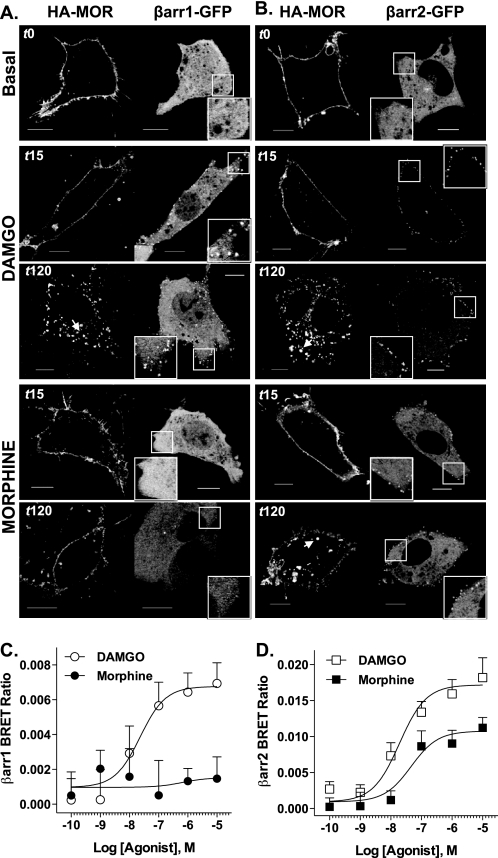
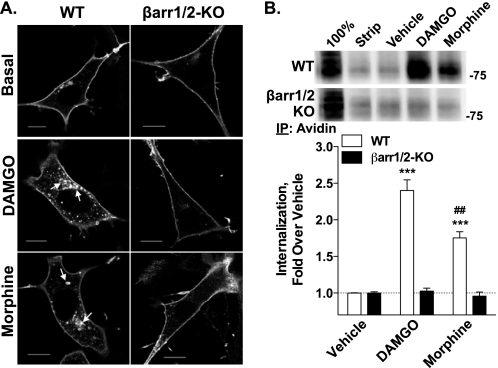
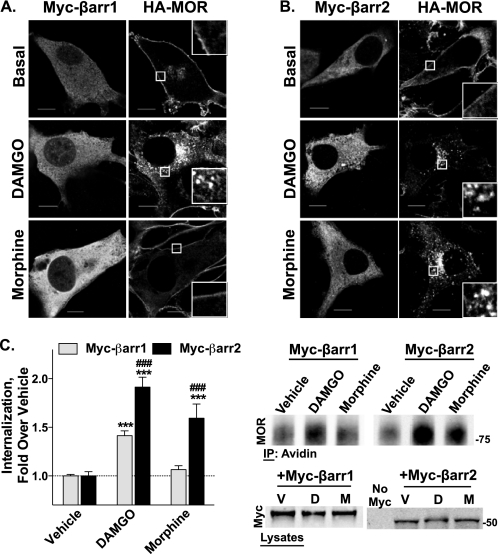
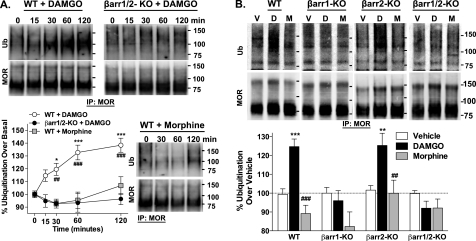
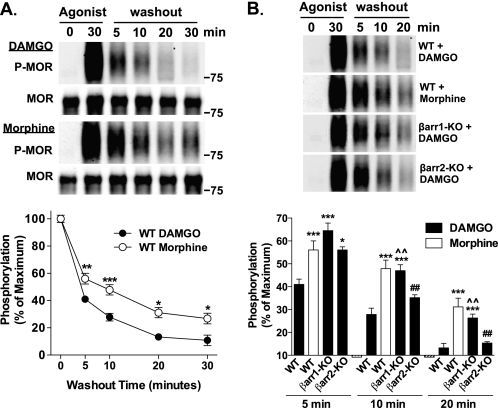
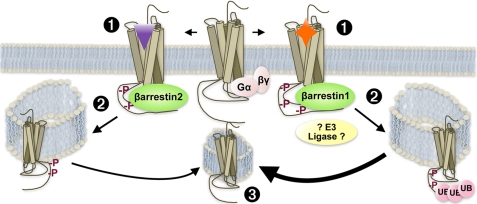
Morphine and other opiates mediate their effects through activation of the μ-opioid receptor (MOR), and regulation of the MOR has been shown to critically affect receptor responsiveness. Activation of the MOR results in receptor phosphorylation, β-arrestin recruitment, and internalization. This classical regulatory process can differ, depending on the ligand occupying the receptor. There are two forms of β-arrestin, β-arrestin1 and β-arrestin2 (also known as arrestin2 and arrestin3, respectively); however, most studies have focused on the consequences of recruiting β-arrestin2 specifically. In this study, we examine the different contributions of β-arrestin1- and β-arrestin2-mediated regulation of the MOR by comparing MOR agonists in cells that lack expression of individual or both β-arrestins. Here we show that morphine only recruits β-arrestin2, whereas the MOR-selective enkephalin [D-Ala(2),N-Me-Phe(4),Gly(5)-ol]enkephalin (DAMGO), recruits either β-arrestin. We show that β-arrestins are required for receptor internalization and that only β-arrestin2 can rescue morphine-induced MOR internalization, whereas either β-arrestin can rescue DAMGO-induced MOR internalization. DAMGO activation of the receptor promotes MOR ubiquitination over time. Interestingly, β-arrestin1 proves to be critical for MOR ubiquitination as modification does not occur in the absence of β-arrestin1 nor when morphine occupies the receptor. Moreover, the selective interactions between the MOR and β-arrestin1 facilitate receptor dephosphorylation, which may play a role in the resensitization of the MOR and thereby contribute to overall development of opioid tolerance.
Figures
References
-
- Bohn L. M., Dykstra L. A., Lefkowitz R. J., Caron M. G., Barak L. S. (2004) Mol. Pharmacol. 66, 106–112 - PubMed
-
- Girnita L., Shenoy S. K., Sehat B., Vasilcanu R., Girnita A., Lefkowitz R. J., Larsson O. (2005) J. Biol. Chem. 280, 24412–24419 - PubMed
-
- Shenoy S. K., McDonald P. H., Kohout T. A., Lefkowitz R. J. (2001) Science 294, 1307–1313 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
