

Cholangiocyte N-Ras protein mediates lipopolysaccharide-induced interleukin 6 secretion and proliferation
- PMID: 21757746
- PMCID: PMC3162394
- DOI: 10.1074/jbc.M111.269464
Cholangiocyte N-Ras protein mediates lipopolysaccharide-induced interleukin 6 secretion and proliferation
Abstract
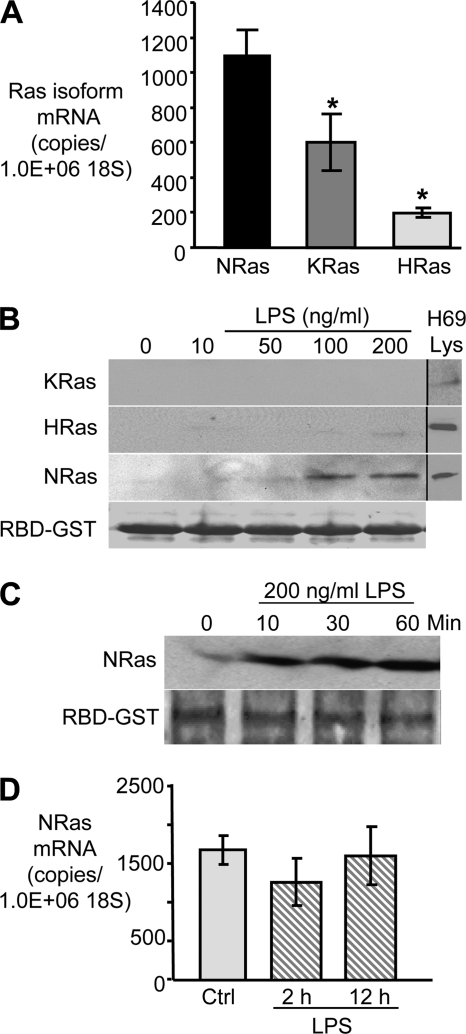
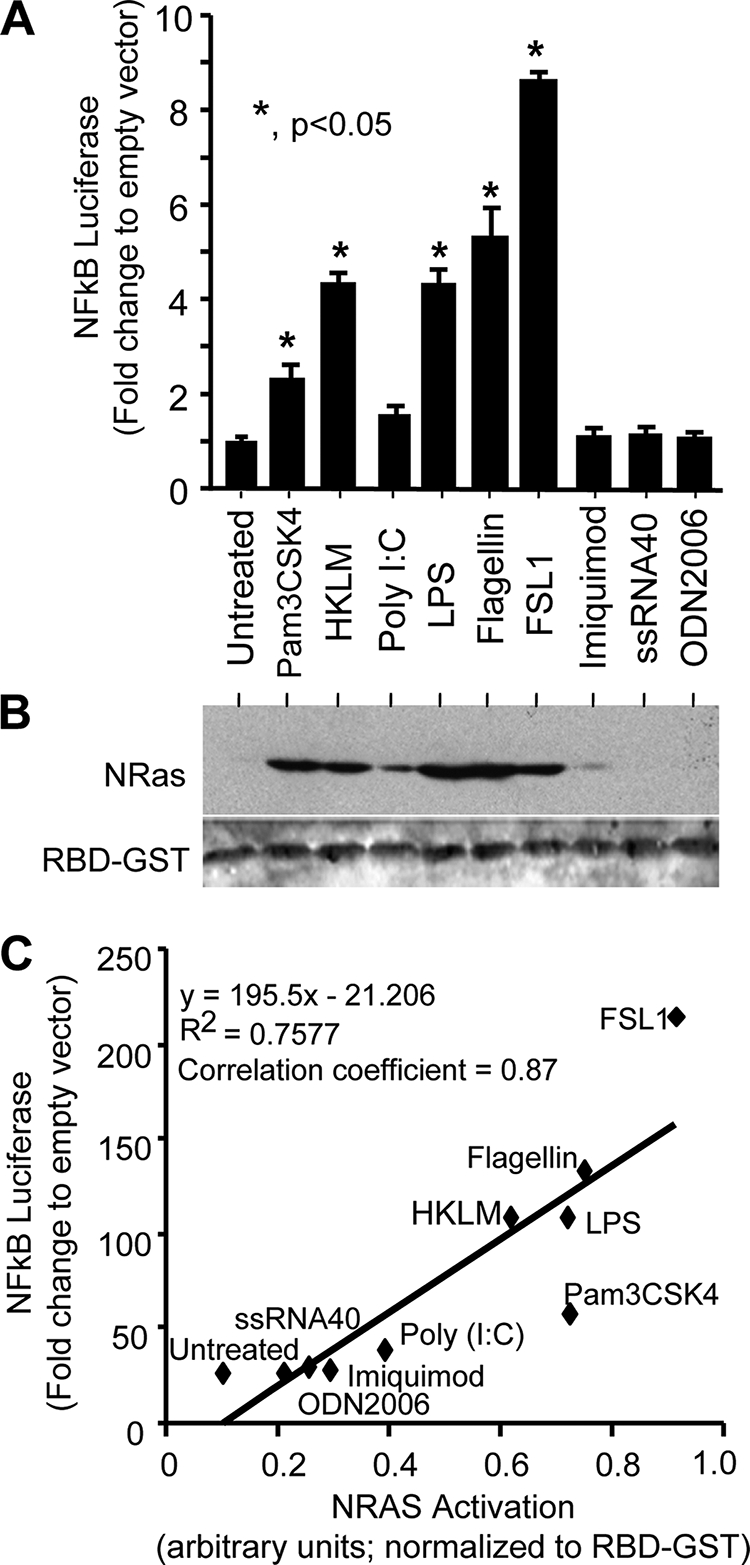
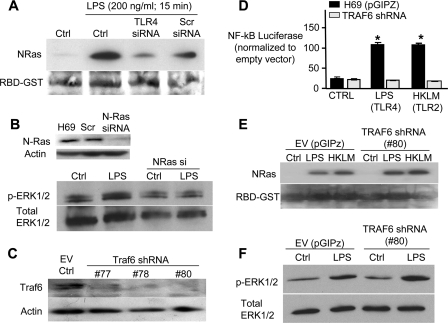
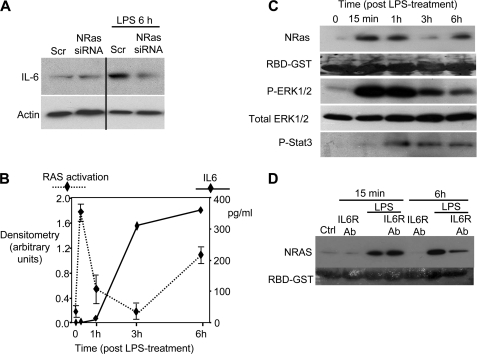
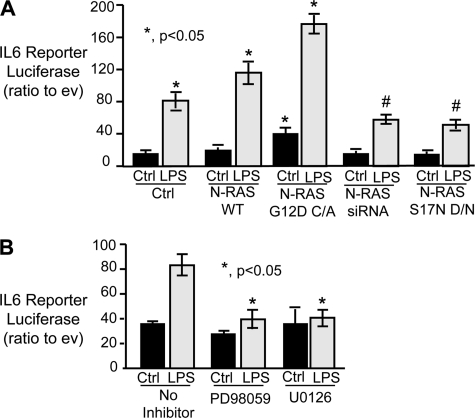
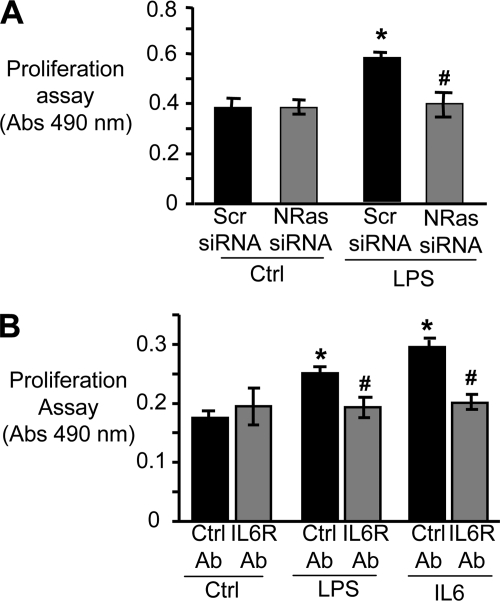
Cholangiocytes, the epithelial cells lining the bile ducts in the liver, are periodically exposed to potentially injurious microbes and/or microbial products. As a result, cholangiocytes actively participate in microbe-associated, hepatic proinflammatory responses. We previously showed that infection of cultured human cholangiocytes with the protozoan parasite, Cryptosporidium parvum, or treatment with gram-negative bacteria-derived LPS, activates NFκB in a myeloid differentiation 88 (MyD88)-dependent manner. Here, we describe a novel signaling pathway initiated by Toll-like receptors (TLRs) involving the small GTPase, Ras, that mediates cholangiocyte proinflammatory cytokine production and induction of cholangiocyte proliferation. Using cultured human cholangiocytes and a Ras activation assay, we found that agonists of plasma membrane TLRs (TLR 1, 2, 4, 5, and 6) rapidly (<10 min) activated N-Ras, but not other p21 Ras isoforms, resulting in the rapid (<15 min) phosphorylation of the downstream Ras effector, ERK1/2. RNA interference-induced depletion of TRAF6, a downstream effector of MyD88 and known activator of MAPK signaling, had no effect on N-Ras activation. Following N-Ras activation the proinflammatory cytokine, IL6, is rapidly secreted. Using a luciferase reporter, we demonstrated that LPS treatment induced IL6 promoter-driven luciferase which was suppressed using MEK/ERK pharmacologic inhibitors (PD98059 or U0126) and RNAi-induced depletion of N-Ras. Finally, we showed that LPS increased cholangiocyte proliferation (1.5-fold), which was inhibited by depletion of N-Ras; TLR agonist-induced proliferation was also inhibited following pretreatment with an IL6 receptor-blocking antibody. Together, our results support a novel signaling axis involving microbial activation of N-Ras likely involved in the cholangiocyte pathogen-induced proinflammatory response.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
