

Pattern-dependent response modulations in motion-sensitive visual interneurons--a model study
- PMID: 21760894
- PMCID: PMC3132178
- DOI: 10.1371/journal.pone.0021488
Pattern-dependent response modulations in motion-sensitive visual interneurons--a model study
Abstract
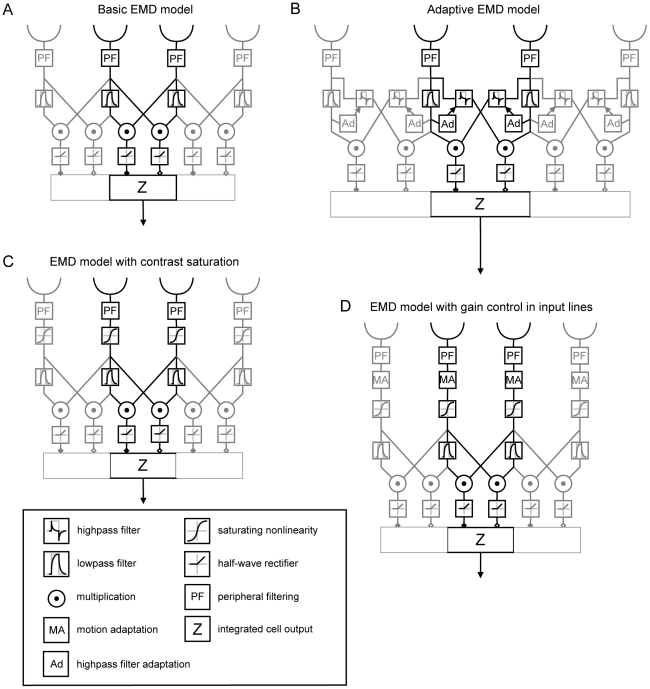
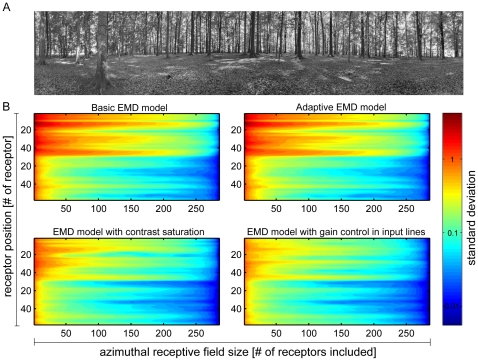
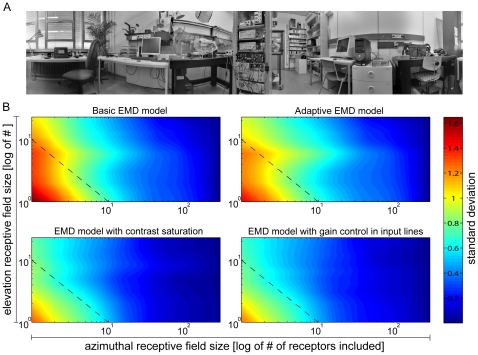
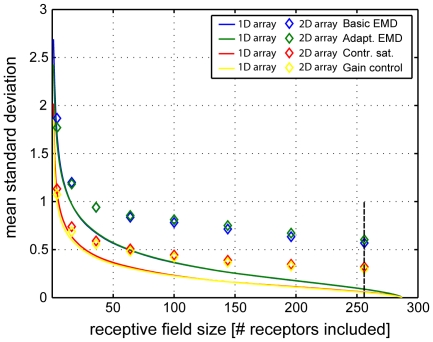
Even if a stimulus pattern moves at a constant velocity across the receptive field of motion-sensitive neurons, such as lobula plate tangential cells (LPTCs) of flies, the response amplitude modulates over time. The amplitude of these response modulations is related to local pattern properties of the moving retinal image. On the one hand, pattern-dependent response modulations have previously been interpreted as 'pattern-noise', because they deteriorate the neuron's ability to provide unambiguous velocity information. On the other hand, these modulations might also provide the system with valuable information about the textural properties of the environment. We analyzed the influence of the size and shape of receptive fields by simulations of four versions of LPTC models consisting of arrays of elementary motion detectors of the correlation type (EMDs). These models have previously been suggested to account for many aspects of LPTC response properties. Pattern-dependent response modulations decrease with an increasing number of EMDs included in the receptive field of the LPTC models, since spatial changes within the visual field are smoothed out by the summation of spatially displaced EMD responses. This effect depends on the shape of the receptive field, being the more pronounced--for a given total size--the more elongated the receptive field is along the direction of motion. Large elongated receptive fields improve the quality of velocity signals. However, if motion signals need to be localized the velocity coding is only poor but the signal provides--potentially useful--local pattern information. These modelling results suggest that motion vision by correlation type movement detectors is subject to uncertainty: you cannot obtain both an unambiguous and a localized velocity signal from the output of a single cell. Hence, the size and shape of receptive fields of motion sensitive neurons should be matched to their potential computational task.
Conflict of interest statement
Figures


References
-
- Lappe M. Neuronal Processing of Optic Flow. Elsevier. 1999;44
-
- Egelhaaf M. The neural computation of visual motion information. Invertebrate Vision. 2006. pp. 399–461.
-
- Reichardt W. Autocorrelation, a principle for the evaluation of sensory information by the central nervous system. In: Rosenblith WA, editor. Sensory Communication. M.I.T. Press; Wiley, J.& Sons; 1961. pp. 303–317.
-
- Egelhaaf M, Borst A, Reichardt W. Computational structure of a biological motion-detection system as revealed by local detector analysis in the fly's nervous system. Journal of the Optical Society of America A: Optics and Image Science. 1989;6:1070–1087. - PubMed
-
- Egelhaaf M, Borst A. Movement detection in arthropods. Reviews of Oculomotor Research. 1993;5:53–77. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
