

V-ATPase-dependent ectodermal voltage and pH regionalization are required for craniofacial morphogenesis
- PMID: 21761475
- PMCID: PMC10277013
- DOI: 10.1002/dvdy.22685
V-ATPase-dependent ectodermal voltage and pH regionalization are required for craniofacial morphogenesis
Abstract
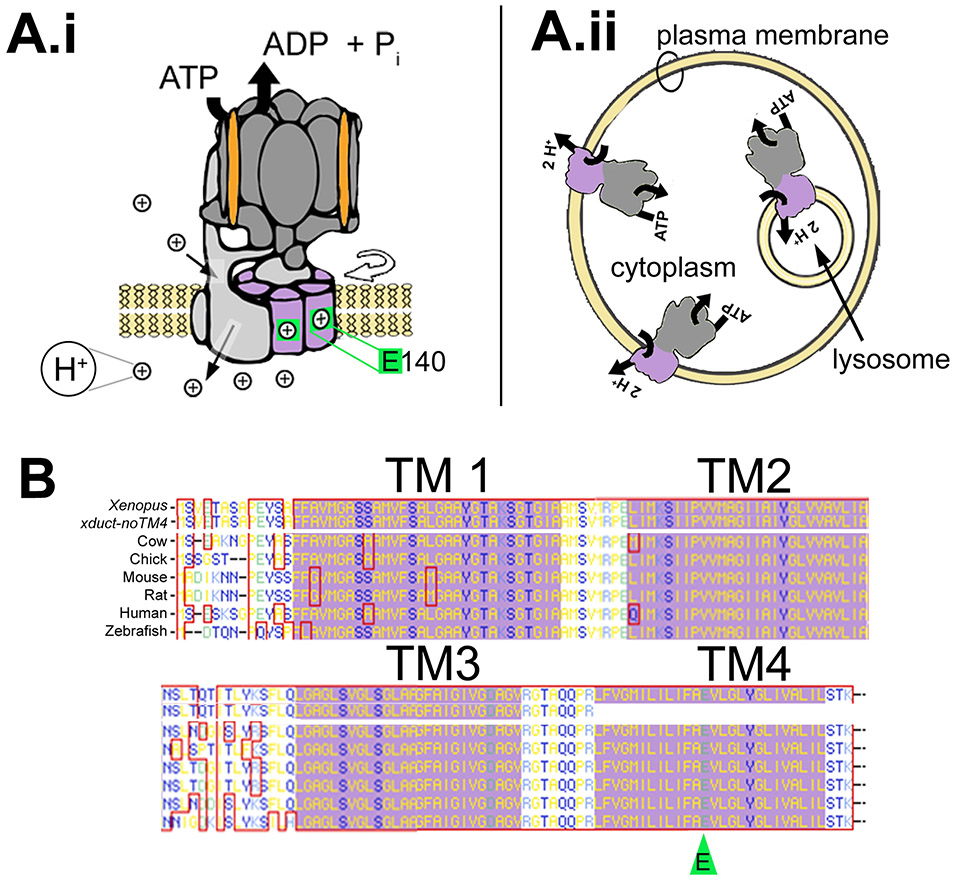
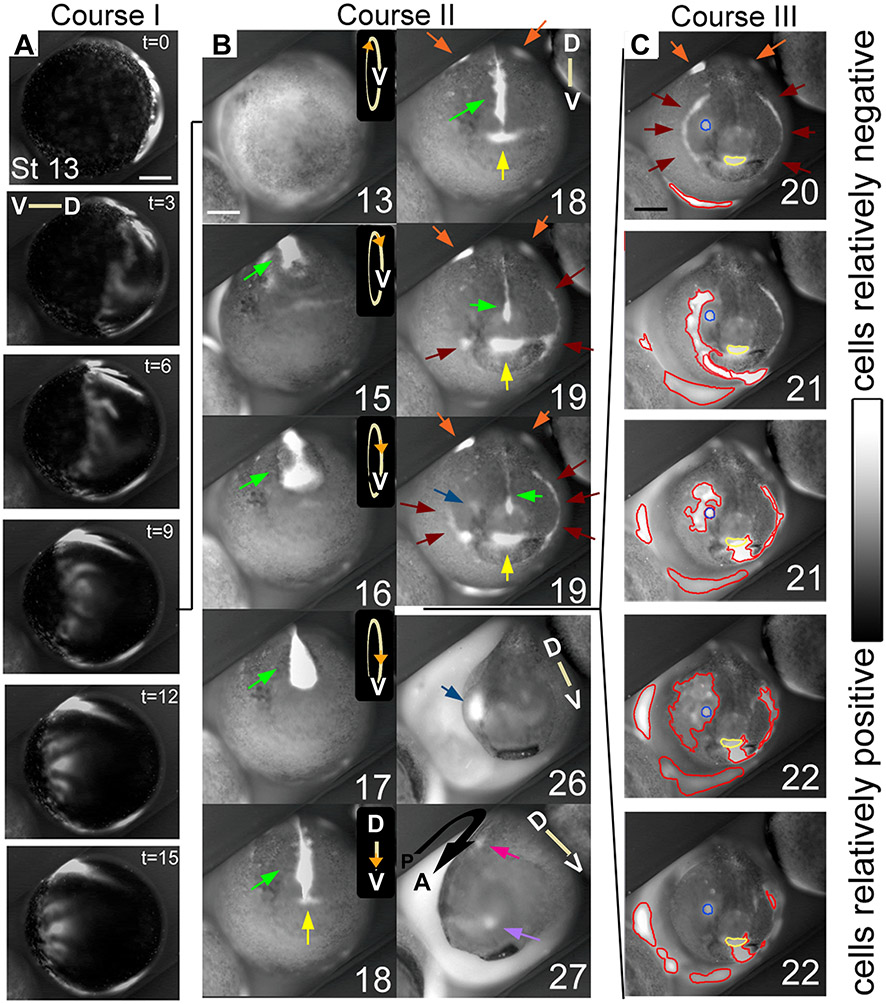
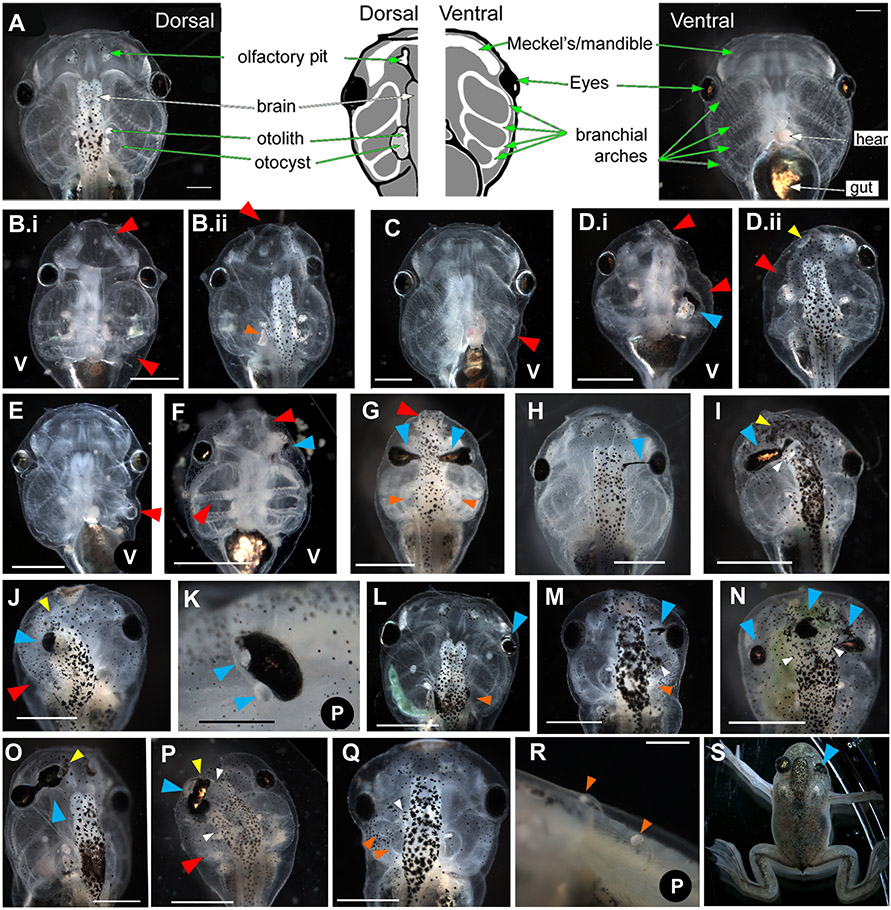
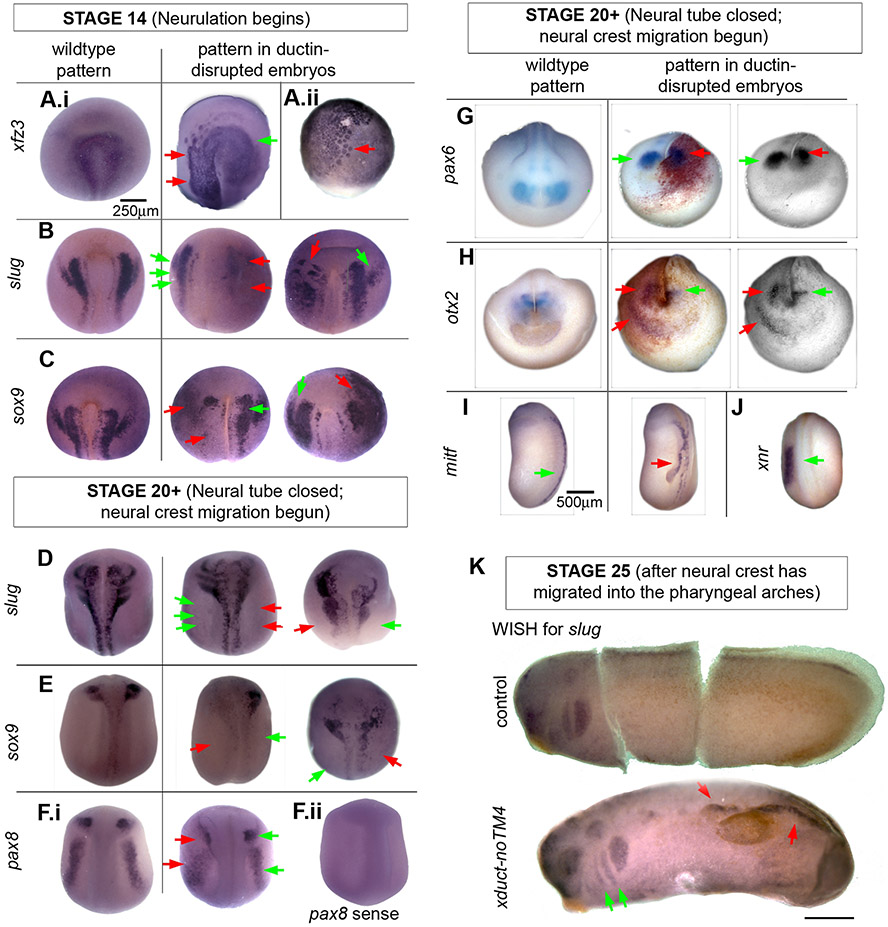
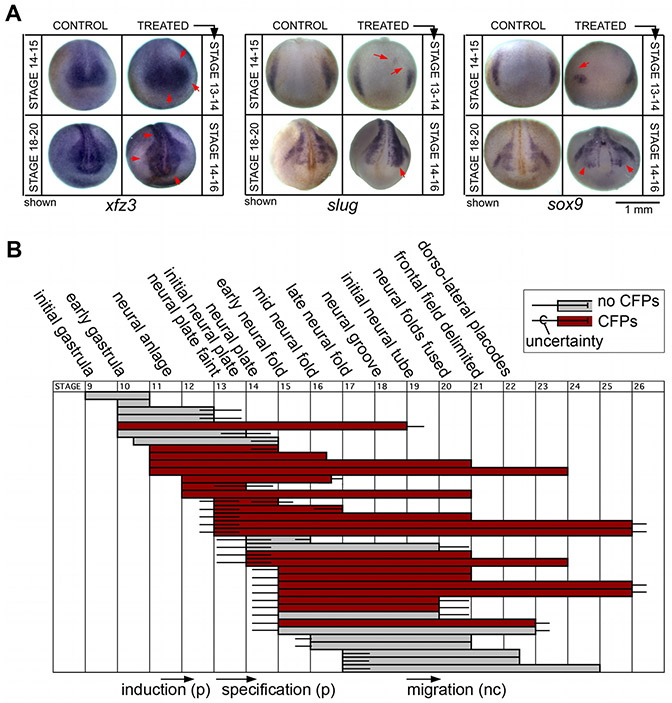
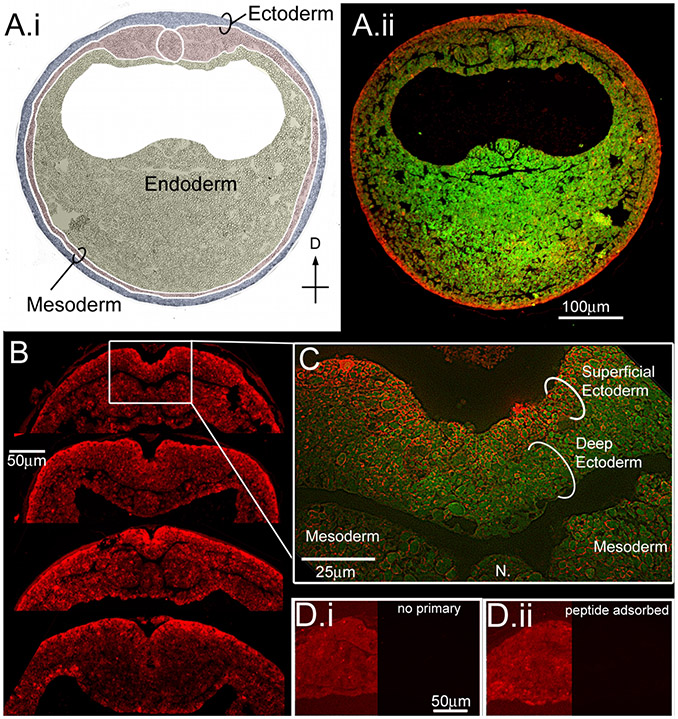
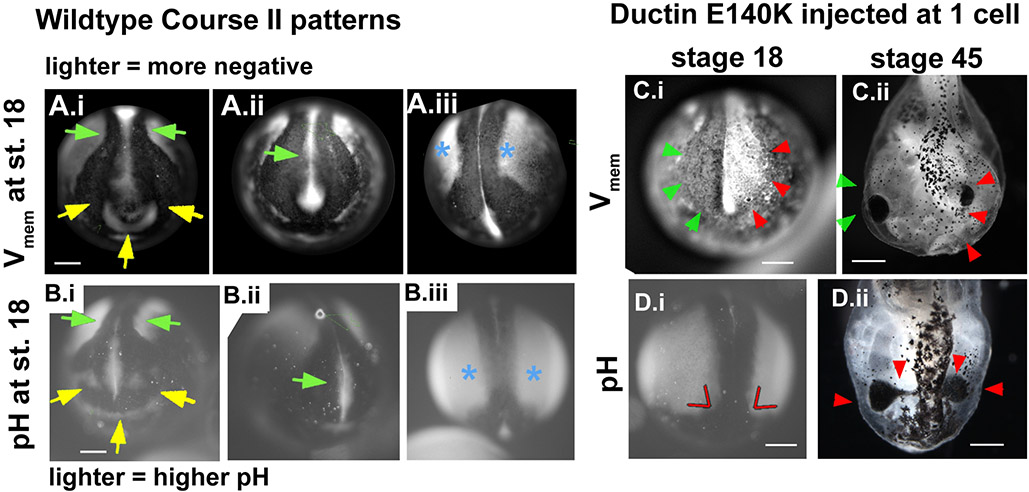
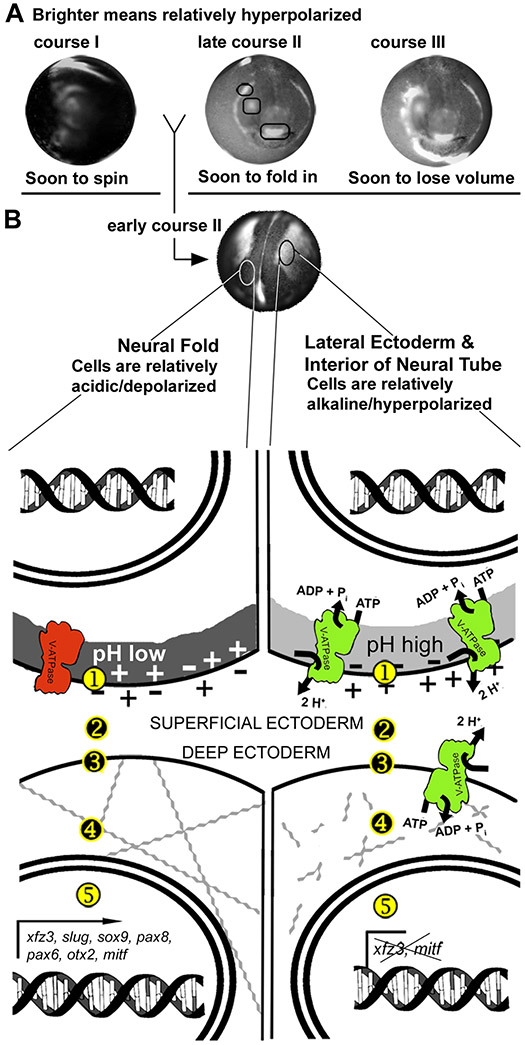
Using voltage and pH reporter dyes, we have discovered a never-before-seen regionalization of the Xenopus ectoderm, with cell subpopulations delimited by different membrane voltage and pH. We distinguished three courses of bioelectrical activity. Course I is a wave of hyperpolarization that travels across the gastrula. Course II comprises the appearance of patterns that match shape changes and gene expression domains of the developing face; hyperpolarization marks folding epithelium and both hyperpolarized and depolarized regions overlap domains of head patterning genes. In Course III, localized regions of hyperpolarization form at various positions, expand, and disappear. Inhibiting H(+) -transport by the H(+) -V-ATPase causes abnormalities in: (1) the morphology of craniofacial structures; (2) Course II voltage patterns; and (3) patterns of sox9, pax8, slug, mitf, xfz3, otx2, and pax6. We conclude that this bioelectric signal has a role in development of the face. Thus, it exemplifies an important, under-studied mechanism of developmental regulation.
Copyright © 2011 Wiley-Liss, Inc.
Figures
References
-
- Adams DS. 2008. A new tool for tissue engineers: ions as regulators of morphogenesis during development and regeneration. Tissue Eng Part A 14:1461–1468. - PubMed
-
- Adams DS, Masi A, Levin M. 2007. H+ pump-dependent changes in membrane voltage are an early mechanism necessary and sufficient to induce Xenopus tail regeneration. Development 134:1323–1335. - PubMed
-
- Anderson T, Sparkowski J, Goldstein DJ, Schlegel R. 1995. Vacuolar H(+)-ATPase mutants transform cells and define a binding site for the papillomavirus E5 oncoprotein. J Biol Chem 270:6830–6837. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
