

Instability of toxin A subunit of AB(5) toxins in the bacterial periplasm caused by deficiency of their cognate B subunits
- PMID: 21762677
- PMCID: PMC5007120
- DOI: 10.1016/j.bbamem.2011.06.016
Instability of toxin A subunit of AB(5) toxins in the bacterial periplasm caused by deficiency of their cognate B subunits
Abstract
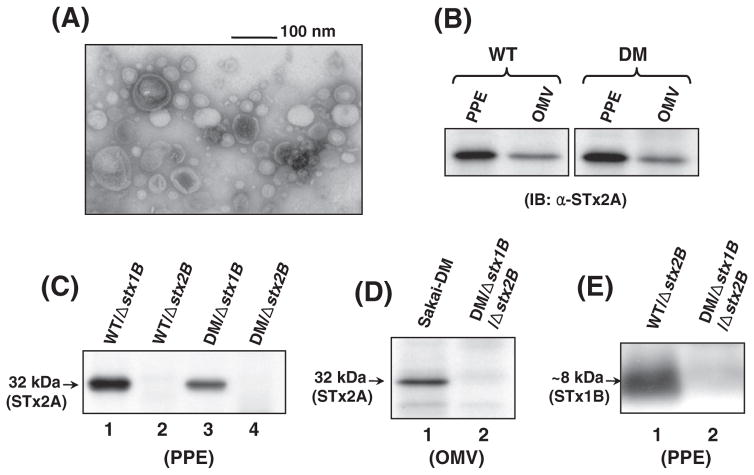
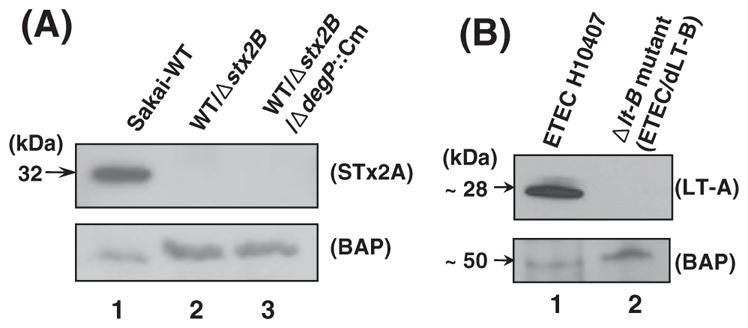
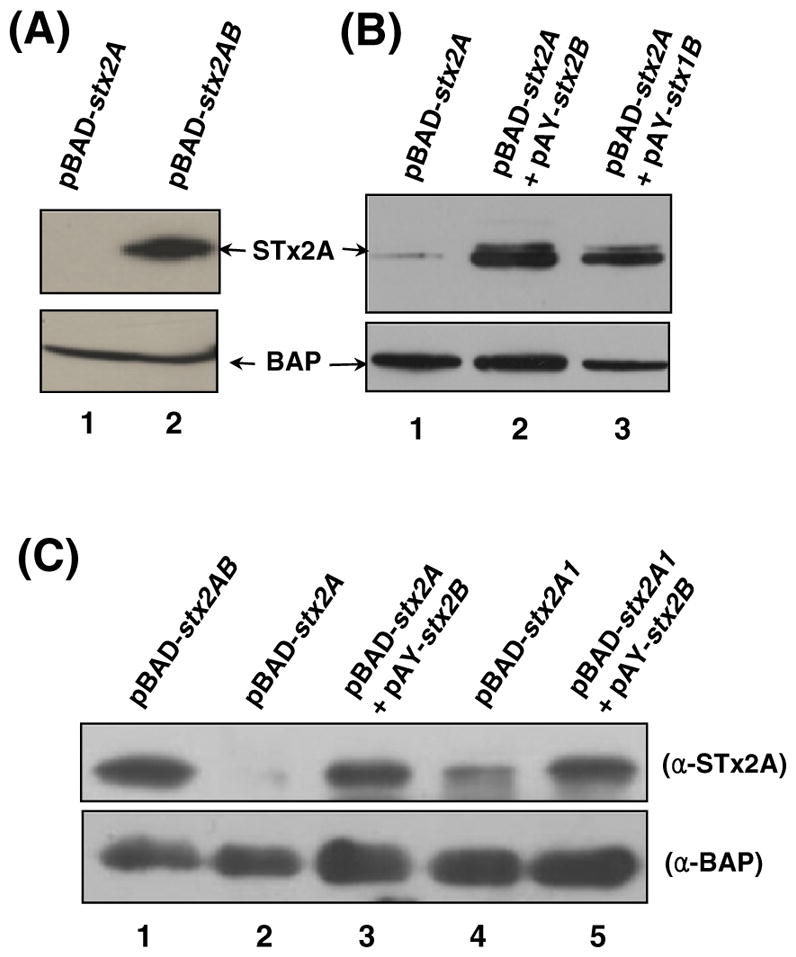
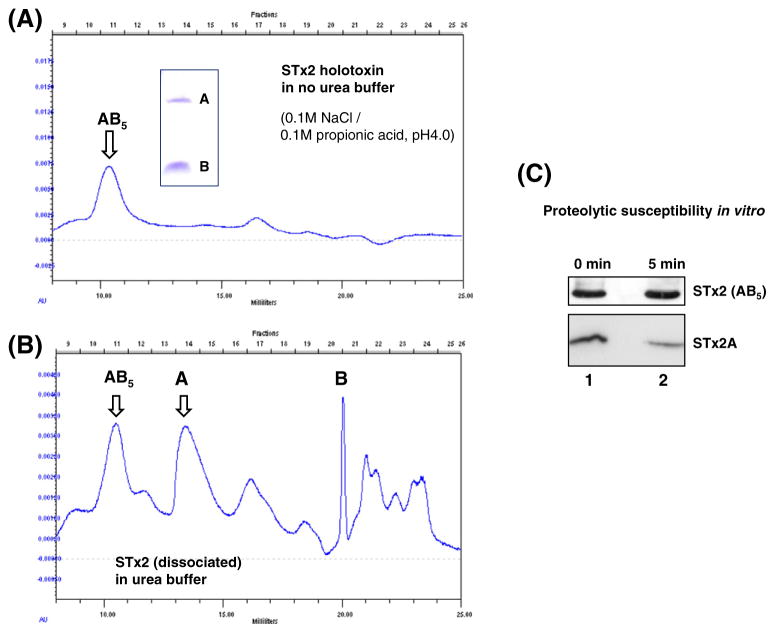
Shiga toxin (STx) belongs to the AB(5) toxin family and is transiently localized in the periplasm before secretion into the extracellular milieu. While producing outer membrane vesicles (OMVs) containing only A subunit of the toxin (STxA), we created specific STx1B- and STx2B-deficient mutants of E. coli O157:H7. Surprisingly, STxA subunit was absent in the OMVs and periplasm of the STxB-deficient mutants. In parallel, the A subunit of heat-labile toxin (LT) of enterotoxigenic E. coli (ETEC) was absent in the periplasm of the LT-B-deficient mutant, suggesting that instability of toxin A subunit in the absence of the B subunit is a common phenomenon in the AB(5) bacterial toxins. Moreover, STx2A was barely detectable in the periplasm of E. coli JM109 when stx2A was overexpressed alone, while it was stably present when stxB was co-expressed. Compared with STx2 holotoxin, purified STx2A was degraded rapidly by periplasmic proteases when assessed for in vitro proteolytic susceptibility, suggesting that the B subunit contributes to stability of the toxin A subunit in the periplasm. We propose a novel role for toxin B subunits of AB(5) toxins in protection of the A subunit from proteolysis during holotoxin assembly in the periplasm.
Copyright © 2011 Elsevier B.V. All rights reserved.
Figures
Similar articles
-
Shiga toxin A subunit mutant of Escherichia coli O157:H7 releases outer membrane vesicles containing the B-pentameric complex.FEMS Immunol Med Microbiol. 2010 Apr;58(3):412-20. doi: 10.1111/j.1574-695X.2010.00654.x. Epub 2010 Jan 22. FEMS Immunol Med Microbiol. 2010. PMID: 20199568
-
The serine 31 residue of the B subunit of Shiga toxin 2 is essential for secretion in enterohemorrhagic Escherichia coli.Infect Immun. 2007 May;75(5):2189-200. doi: 10.1128/IAI.01546-06. Epub 2007 Feb 26. Infect Immun. 2007. PMID: 17325057 Free PMC article.
-
Excess A-subunits of Shiga toxin 2a are produced in enterohemorrhagic Escherichia coli.Sci Rep. 2025 May 14;15(1):16712. doi: 10.1038/s41598-025-01342-2. Sci Rep. 2025. PMID: 40368985 Free PMC article.
-
Vaccines for viral and bacterial pathogens causing acute gastroenteritis: Part II: Vaccines for Shigella, Salmonella, enterotoxigenic E. coli (ETEC) enterohemorragic E. coli (EHEC) and Campylobacter jejuni.Hum Vaccin Immunother. 2015;11(3):601-19. doi: 10.1080/21645515.2015.1011578. Hum Vaccin Immunother. 2015. PMID: 25715096 Free PMC article. Review.
-
Shiga/verocytotoxins and Shiga/verotoxigenic Escherichia coli in animals.Vet Res. 1999 Mar-Jun;30(2-3):235-57. Vet Res. 1999. PMID: 10367357 Review.
Cited by
-
Molecular Biology of Escherichia Coli Shiga Toxins' Effects on Mammalian Cells.Toxins (Basel). 2020 May 23;12(5):345. doi: 10.3390/toxins12050345. Toxins (Basel). 2020. PMID: 32456125 Free PMC article. Review.
-
A Shiga Toxin B-Subunit-Based Lectibody Boosts T Cell Cytotoxicity towards Gb3-Positive Cancer Cells.Cells. 2023 Jul 20;12(14):1896. doi: 10.3390/cells12141896. Cells. 2023. PMID: 37508560 Free PMC article.
-
Different roles of the C-terminal end of Stx1A and Stx2A for AB5 complex integrity and retrograde transport of Stx in HeLa cells.Pathog Dis. 2015 Dec;73(9):ftv083. doi: 10.1093/femspd/ftv083. Epub 2015 Oct 5. Pathog Dis. 2015. PMID: 26443836 Free PMC article.
-
A Conformational Shift in the Dissociated Cholera Toxin A1 Subunit Prevents Reassembly of the Cholera Holotoxin.Toxins (Basel). 2015 Jul 20;7(7):2674-84. doi: 10.3390/toxins7072674. Toxins (Basel). 2015. PMID: 26266549 Free PMC article.
-
Proline Isomerization as a Key Determinant for Hsp90-Toxin Interactions.Front Cell Infect Microbiol. 2021 Oct 22;11:771653. doi: 10.3389/fcimb.2021.771653. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 34746036 Free PMC article.
References
-
- Fraser ME, Fujinaga M, Cherney MM, et al. Structure of Shiga toxin type 2 (Stx2) from Escherichia coli O157:H7. J Biol Chem. 2004;279:27511–27517. - PubMed
-
- Johannes L, Römer W. Shiga toxins — from cell biology to biomedical applications. Nat Rev Microbiol. 2010;8:105–116. - PubMed
-
- Endo Y, Tsurugi K, Yutsudo T, et al. Site of action of a Vero toxin (VT2) from Escherichia coli O157:H7 and of Shiga toxin on eukaryotic ribosomes. RNA N-glycosidase activity of the toxins. Eur J Biochem. 1988;171:45–50. - PubMed
-
- Lingwood CA. Role of verotoxin receptors in pathogenesis. Trends Microbiol. 1996;4:147–153. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
