

Duct cells contribute to regeneration of endocrine and acinar cells following pancreatic damage in adult mice
- PMID: 21763240
- PMCID: PMC4326039
- DOI: 10.1053/j.gastro.2011.07.003
Duct cells contribute to regeneration of endocrine and acinar cells following pancreatic damage in adult mice
Abstract
Background & aims: There have been conflicting results on a cell of origin in pancreatic regeneration. These discrepancies predominantly stem from lack of specific markers for the pancreatic precursors/stem cells, as well as differences in the targeted cells and severity of tissue injury in the experimental models so far proposed. We attempted to create a model that used diphtheria toxin receptor (DTR) to ablate specific cell populations, control the extent of injury, and avoid induction of the inflammatory response.
Methods: To target specific types of pancreatic cells, we crossed R26DTR or R26DTR/lacZ mice with transgenic mice that express the Cre recombinase in the pancreas, under control of the Pdx1 (global pancreatic) or elastase (acinar-specific) promoters.
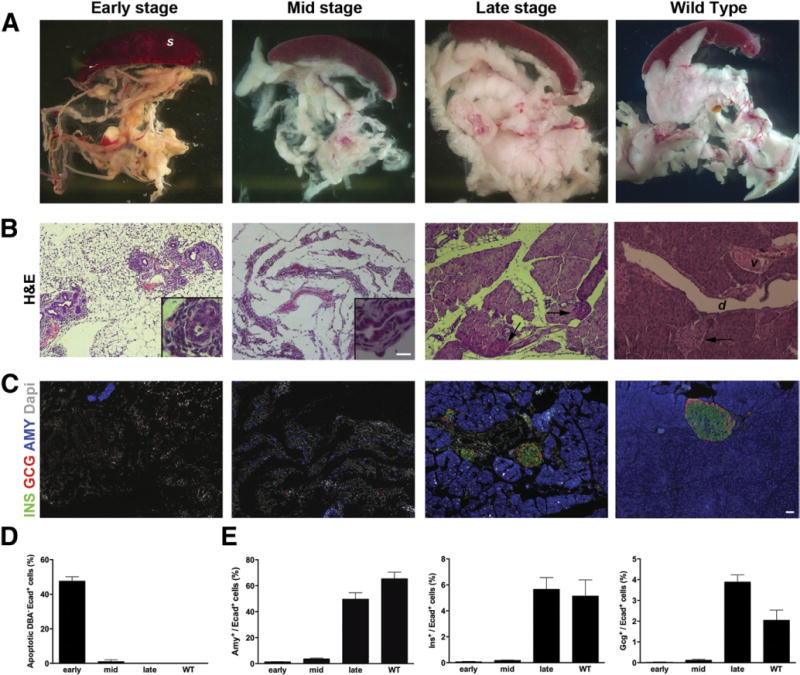
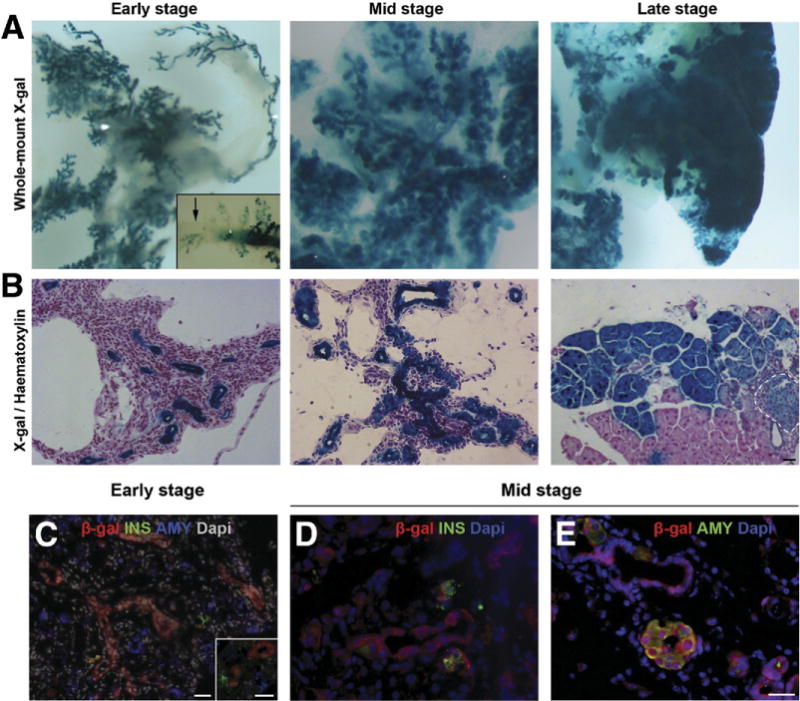
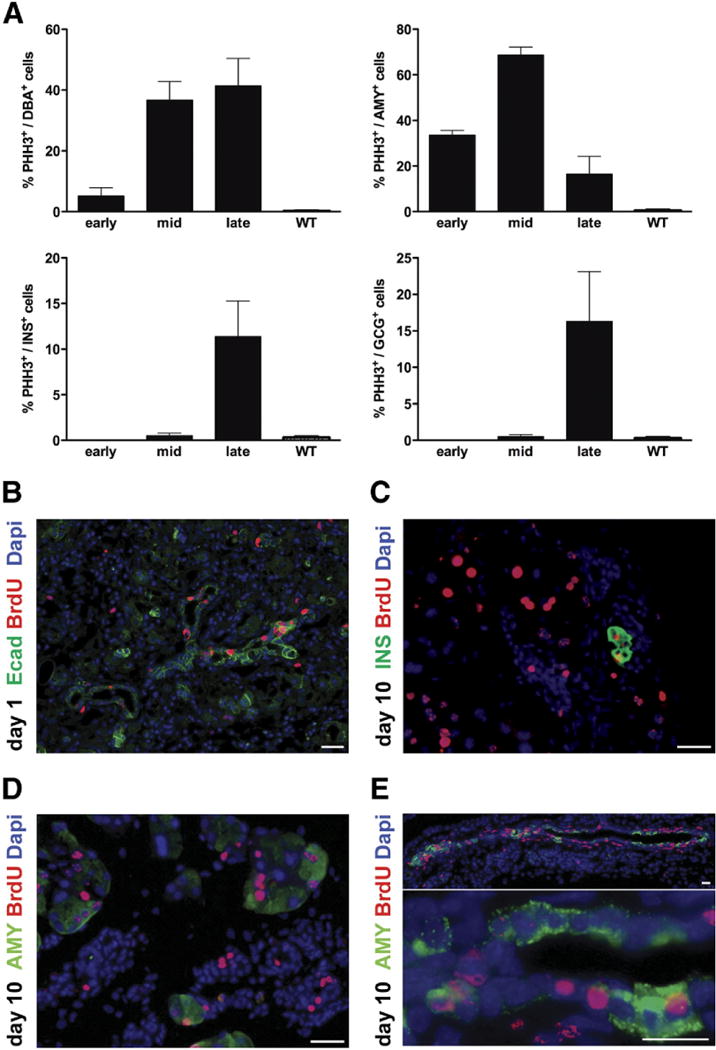
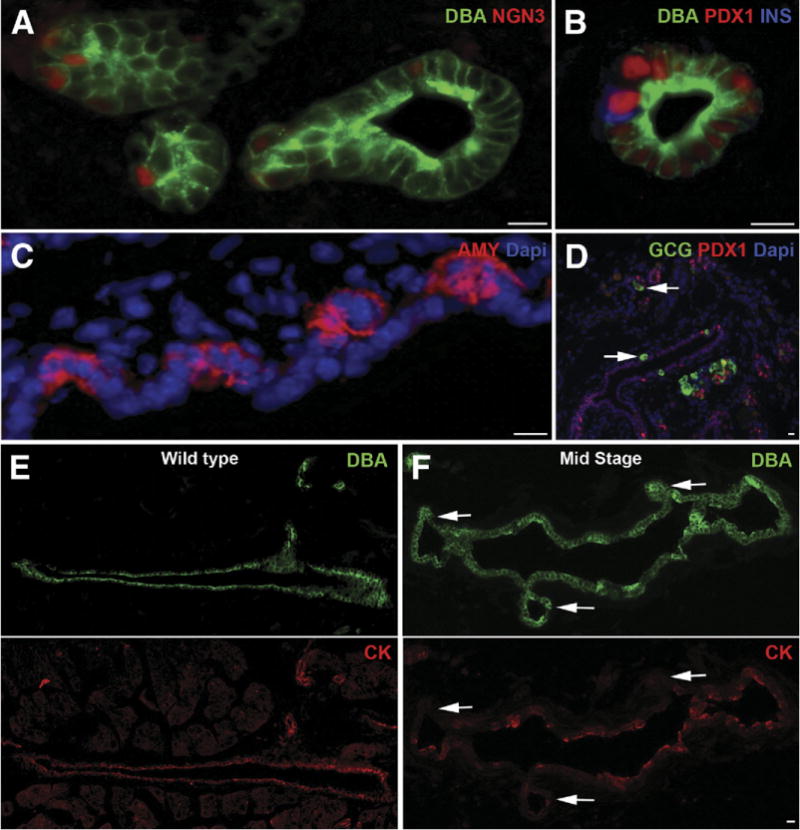
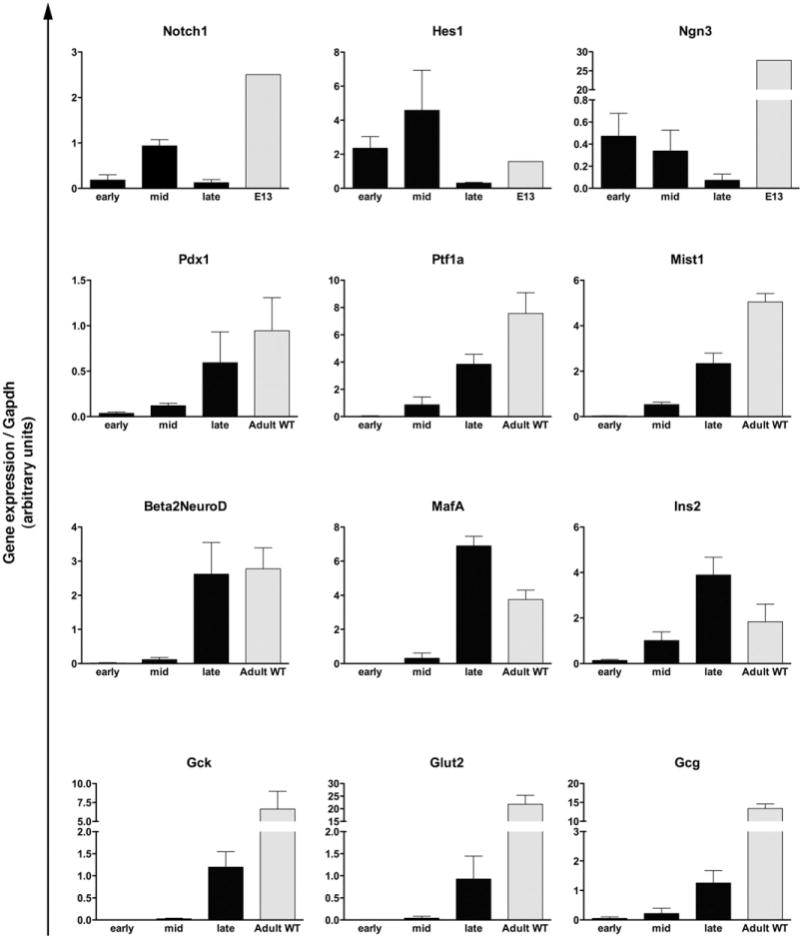
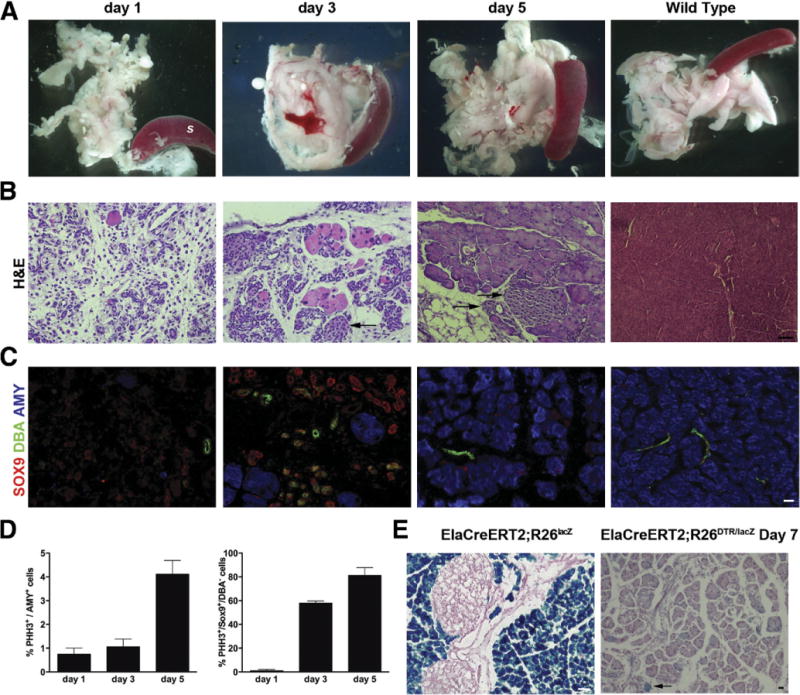
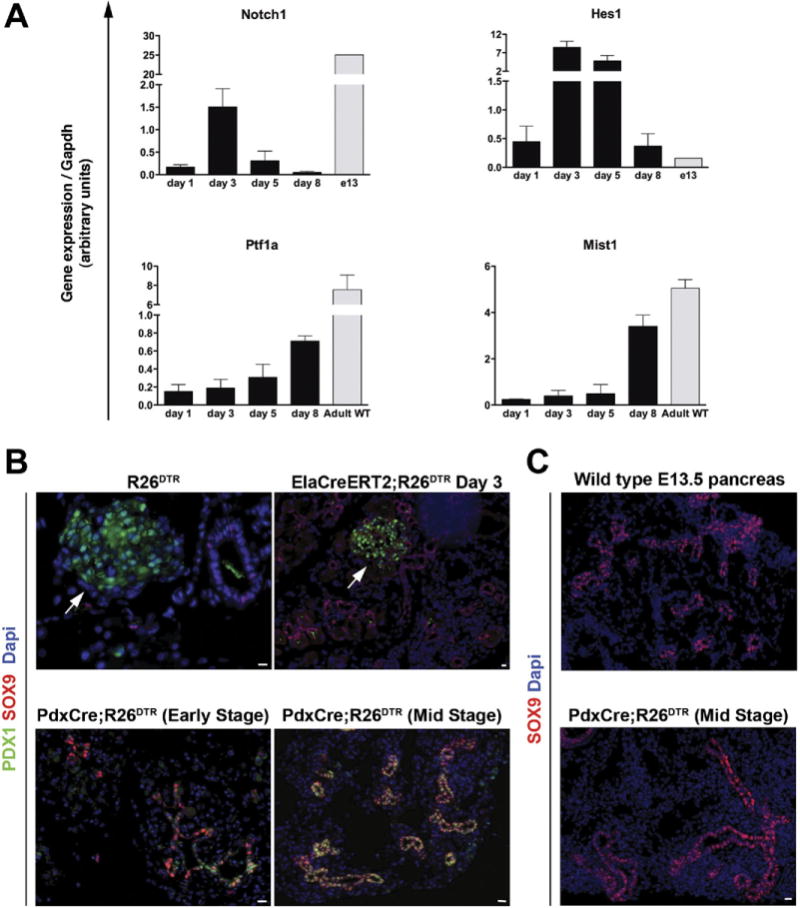
Results: Exposure of PdxCre;R26DTR mice to diphtheria toxin resulted in extensive ablation of acinar and endocrine tissues but not ductal cells. Surviving cells within the ductal compartment contributed to regeneration of endocrine and acinar cells via recapitulation of the embryonic pancreatic developmental program. However, following selective ablation of acinar tissue in ElaCreERT2;R26DTR mice, regeneration likely occurred by reprogramming of ductal cells to acinar lineage.
Conclusions: In the pancreas of adult mice, epithelial cells within the ductal compartment contribute to regeneration of endocrine and acinar cells. The severity of injury determines the regenerative mechanisms and cell types that contribute to this process.
Copyright © 2011 AGA Institute. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflicts of interest
The authors disclose no conflicts.
Figures
References
-
- Dor Y, Brown J, Martinez OI, et al. Adult pancreatic beta-cells are formed by self-duplication rather than stem-cell differentiation. Nature. 2004;429:41–46. - PubMed
-
- Teta M, Rankin MM, Long SY, et al. Growth and regeneration of adult beta cells does not involve specialized progenitors. Dev Cell. 2007;12:817–826. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
