

Multifaceted counter-APOBEC3G mechanisms employed by HIV-1 Vif
- PMID: 21763507
- PMCID: PMC3139112
- DOI: 10.1016/j.jmb.2011.03.058
Multifaceted counter-APOBEC3G mechanisms employed by HIV-1 Vif
Abstract
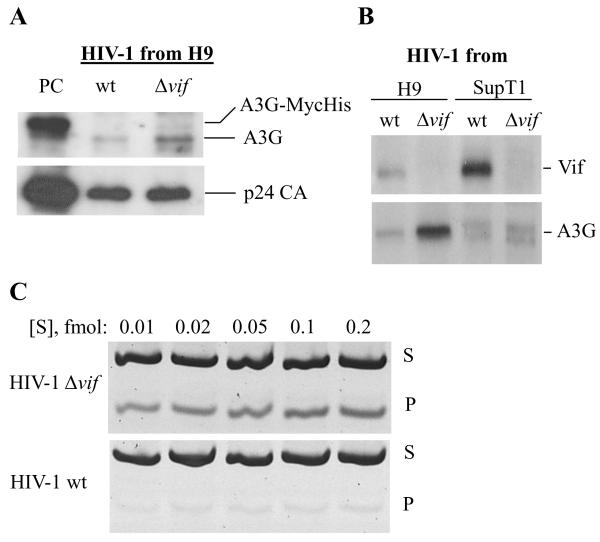
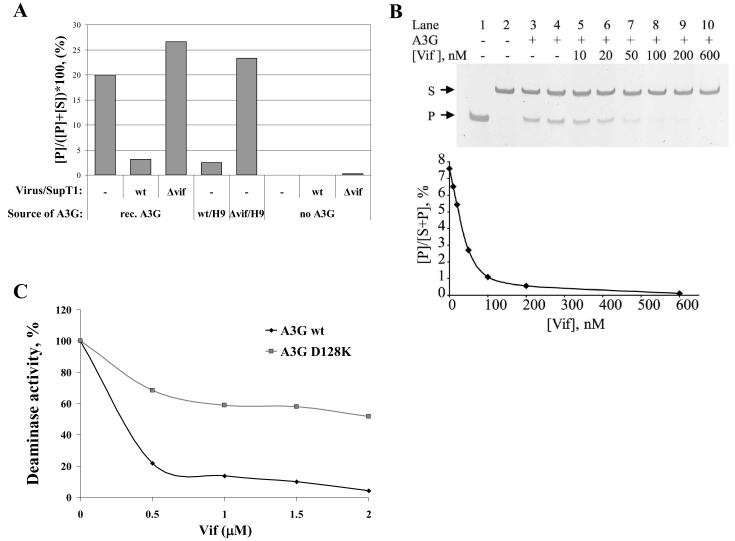
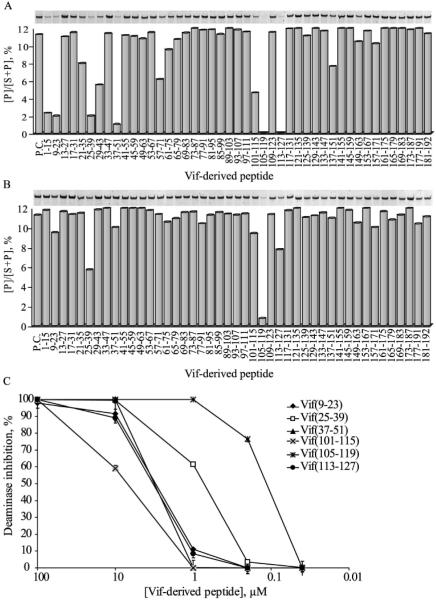
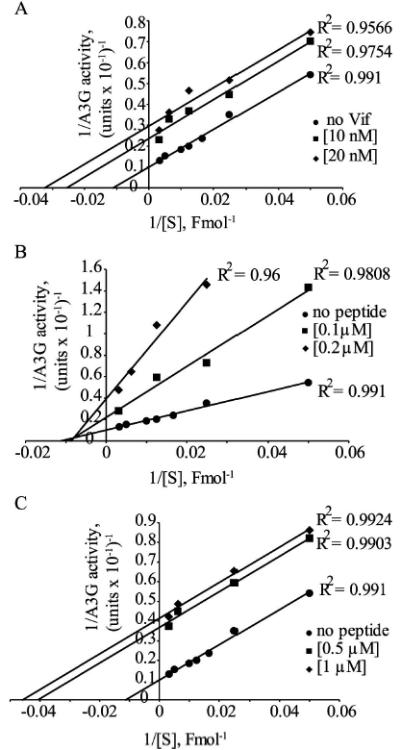
In the absence of human immunodeficiency virus type 1 (HIV-1) Vif protein, the host antiviral deaminase apolipoprotein B mRNA-editing enzyme-catalytic polypeptide-like 3G (A3G) restricts the production of infectious HIV-1 by deamination of dC residues in the negative single-stranded DNA produced by reverse transcription. The Vif protein averts the lethal threat of deamination by precluding the packaging of A3G into assembling virions by mediating proteasomal degradation of A3G. In spite of this robust Vif activity, residual A3G molecules that escape degradation and incorporate into newly assembled virions are potentially deleterious to the virus. We hypothesized that virion-associated Vif inhibits A3G enzymatic activity and therefore prevents lethal mutagenesis of the newly synthesized viral DNA. Here, we show that (i) Vif-proficient HIV-1 particles released from H9 cells contain A3G with lower specific activity compared with Δvif-virus-associated A3G, (ii) encapsidated HIV-1 Vif inhibits the deamination activity of recombinant A3G, and (iii) purified HIV-1 Vif protein and the Vif-derived peptide Vif25-39 inhibit A3G activity in vitro at nanomolar concentrations in an uncompetitive manner. Our results manifest the potentiality of Vif to control the deamination threat in virions or in the pre-integration complexes following entry to target cells. Hence, virion-associated Vif could serve as a last line of defense, protecting the virus against A3G antiviral activity.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Characterization of conserved motifs in HIV-1 Vif required for APOBEC3G and APOBEC3F interaction.J Mol Biol. 2008 Sep 12;381(4):1000-11. doi: 10.1016/j.jmb.2008.06.061. Epub 2008 Jun 28. J Mol Biol. 2008. PMID: 18619467
-
HIV-1 Vif inhibits G to A hypermutations catalyzed by virus-encapsidated APOBEC3G to maintain HIV-1 infectivity.Retrovirology. 2014 Oct 11;11:89. doi: 10.1186/s12977-014-0089-5. Retrovirology. 2014. PMID: 25304135 Free PMC article.
-
Identification of a novel HIV-1 inhibitor targeting Vif-dependent degradation of human APOBEC3G protein.J Biol Chem. 2015 Apr 17;290(16):10504-17. doi: 10.1074/jbc.M114.626903. Epub 2015 Feb 27. J Biol Chem. 2015. PMID: 25724652 Free PMC article.
-
Various strategies for developing APOBEC3G protectors to circumvent human immunodeficiency virus type 1.Eur J Med Chem. 2023 Mar 15;250:115188. doi: 10.1016/j.ejmech.2023.115188. Epub 2023 Feb 6. Eur J Med Chem. 2023. PMID: 36773550 Review.
-
[Advances in the study of molecular mechanism of APOBEC3G anti-HIV-1].Yao Xue Xue Bao. 2008 Jul;43(7):678-82. Yao Xue Xue Bao. 2008. PMID: 18819469 Review. Chinese.
Cited by
-
Degradation-Independent Inhibition of APOBEC3G by the HIV-1 Vif Protein.Viruses. 2021 Apr 3;13(4):617. doi: 10.3390/v13040617. Viruses. 2021. PMID: 33916704 Free PMC article. Review.
-
Running loose or getting lost: how HIV-1 counters and capitalizes on APOBEC3-induced mutagenesis through its Vif protein.Viruses. 2012 Nov 14;4(11):3132-61. doi: 10.3390/v4113132. Viruses. 2012. PMID: 23202519 Free PMC article. Review.
-
APOBEC Enzymes as Targets for Virus and Cancer Therapy.Cell Chem Biol. 2018 Jan 18;25(1):36-49. doi: 10.1016/j.chembiol.2017.10.007. Epub 2017 Nov 16. Cell Chem Biol. 2018. PMID: 29153851 Free PMC article. Review.
-
Prototype foamy virus Bet impairs the dimerization and cytosolic solubility of human APOBEC3G.J Virol. 2013 Aug;87(16):9030-40. doi: 10.1128/JVI.03385-12. Epub 2013 Jun 12. J Virol. 2013. PMID: 23760237 Free PMC article.
-
Aromatic disulfides as potential inhibitors against interaction between deaminase APOBEC3G and HIV infectivity factor.Acta Biochim Biophys Sin (Shanghai). 2022 May 25;54(5):725-735. doi: 10.3724/abbs.2022049. Acta Biochim Biophys Sin (Shanghai). 2022. PMID: 35920198 Free PMC article.
References
-
- Jarmuz A, Chester A, Bayliss J, Gisbourne J, Dunham I, Scott J, Navaratnam N. An anthropoid-specific locus of orphan C to U RNA-editing enzymes on chromosome 22. Genomics. 2002;79:285–96. - PubMed
-
- Wedekind JE, Dance GS, Sowden MP, Smith HC. Messenger RNA editing in mammals: new members of the APOBEC family seeking roles in the family business. Trends Genet. 2003;19:207–16. - PubMed
-
- Conticello SG, Thomas CJ, Petersen-Mahrt SK, Neuberger MS. Evolution of the AID/APOBEC family of polynucleotide (deoxy)cytidine deaminases. Mol Biol Evol. 2005;22:367–77. - PubMed
-
- Muramatsu M, Sankaranand VS, Anant S, Sugai M, Kinoshita K, Davidson NO, Honjo T. Specific expression of activation-induced cytidine deaminase (AID), a novel member of the RNA-editing deaminase family in germinal center B cells. J Biol Chem. 1999;274:18470–6. - PubMed
-
- Muramatsu M, Kinoshita K, Fagarasan S, Yamada S, Shinkai Y, Honjo T. Class switch recombination and hypermutation require activation-induced cytidine deaminase (AID), a potential RNA editing enzyme. Cell. 2000;102:553–63. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
