

Review
doi: 10.1016/j.devcel.2011.06.001.
Extracellular movement of signaling molecules
Affiliations
- PMID: 21763615
- PMCID: PMC3220056
- DOI: 10.1016/j.devcel.2011.06.001
Item in Clipboard
Review
Extracellular movement of signaling molecules
Dev Cell.
.
Abstract
Extracellular signaling molecules have crucial roles in development and homeostasis, and their incorrect deployment can lead to developmental defects and disease states. Signaling molecules are released from sending cells, travel to target cells, and act over length scales of several orders of magnitude, from morphogen-mediated patterning of small developmental fields to hormonal signaling throughout the organism. We discuss how signals are modified and assembled for transport, which routes they take to reach their targets, and how their range is affected by mobility and stability.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
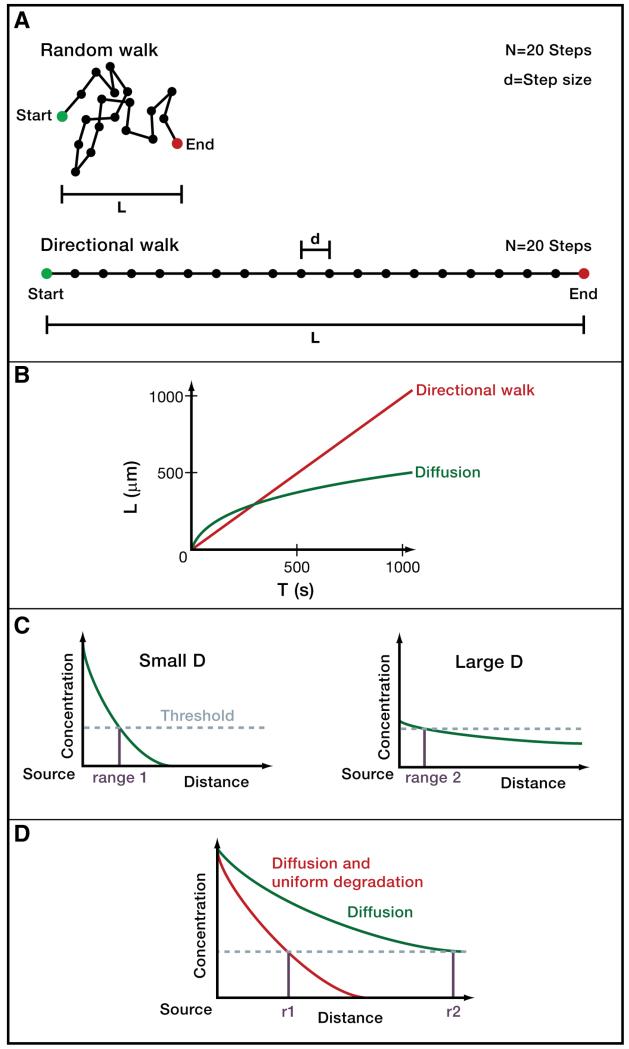
A) Directional movement and random walks. The distance L that a molecule moving in a constant direction covers with N steps of step size d is L= Nd. However, diffusing molecules do not move in a constant direction but rather undergo random walks, in which the direction of motion changes randomly after each step due to collision with surrounding molecules. A diffusing molecule will therefore on average cover a distance (Berg 1993; Phillips et al. 2009). For example, with 20 steps of size 1 a molecule moving in a constant direction could travel a distance L=20, whereas a randomly walking molecule would only be displaced from its starting position by about L=5 on average. Conversely, to travel a distance of L=20, a randomly walking molecule would need to make 400 steps of size 1 on average. B) Timescales of diffusive and directional movement. The displacement L in micrometers as a function of time t (in seconds) for a molecule moving in a constant direction with a velocity v of 1 μm/s is described by L=tv (red). The average displacement L as a function of time t for an ensemble of molecules diffusing with a diffusion coefficient D of 100 μm2/s (e.g. a small protein diffusing in water) is described by (green). Diffusing molecules can move rapidly away from their starting positions over short distances, but take a long time to move long distances. C) Concentration thresholds, signaling range and diffusivity in paracrine signaling. Typically, cells must be exposed to a concentration of paracrine signal above a certain threshold (indicated by dashed horizontal line) in order to respond to signaling. Signals with large diffusion coefficients (right graph) travel farther from their sources than signals with small diffusion coefficients (left graph). However, less diffusive signals can counter-intuitively have longer signaling ranges (r1) than more diffusive signals (r2), since these relatively immobile signals “pile up” near the source at levels above the threshold required for a cellular response. D) Mobility and stability affect signaling range. Molecules that are cleared uniformly and rapidly in the target field accumulate to lower concentrations.
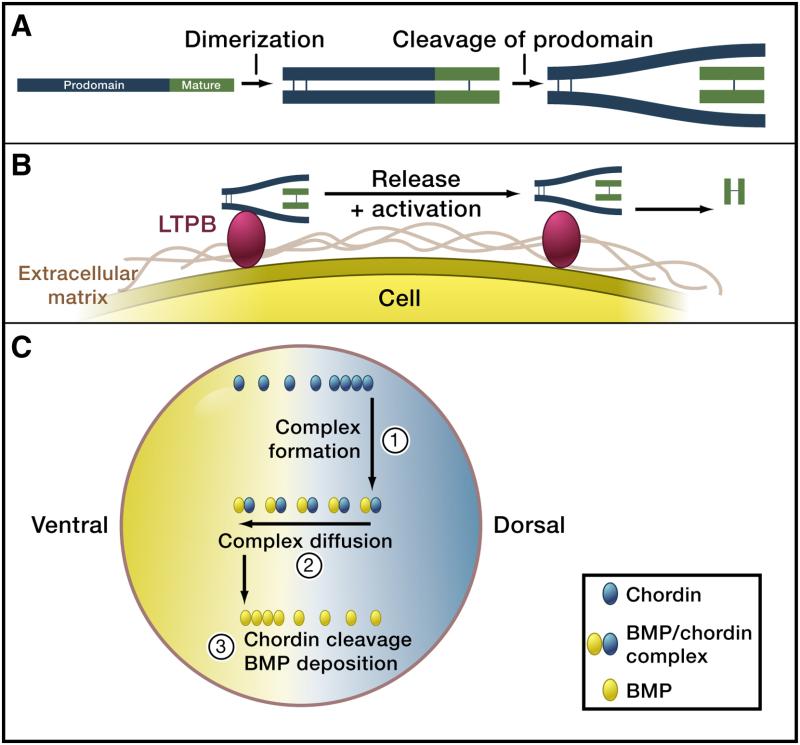
A) Pro-protein cleavage. TGFβ superfamily ligands are produced as pro-proteins, dimerize and require cleavage of the pro-domain by convertases (e.g. Furin). For many TGFβ ligands the prodomain (blue) stays attached to the mature domain (green) after cleavage. Modified from (ten Dijke and Arthur 2007). B) Tethering to the extracellular matrix and release. Pro-domain – mature domain complexes can be tethered to the extracellular matrix (brown), e.g. via the interaction of the pro-domain with latent TGFβ binding proteins (LTBP, red). After cleavage of LTBP and the pro-domain (e.g. by matrix metalloproteinases such as BMP1 and MMP2), the mature domain is released and can now signal to distant cells. Modified from (ten Dijke and Arthur 2007). C) Heteromerization with carrier proteins and shuttling. BMP molecules (green) are thought to be relatively immobile unless bound to Chordin (red). During early embryogenesis in frogs, BMPs are uniformly distributed. Production of Chordin on the dorsal side of the embryo leads to complex formation between BMPs and Chordin. The BMP/Chordin complex is mobile and diffuses. Repeated rounds of Chordin cleavage by a uniformly distributed protease, subsequent release of free BMP and remobilization by Chordin binding is thought to eventually result in the accumulation of free BMP on the ventral side. Additional downstream feedback signaling networks can result in robust pattern formation. Figure modified from (Lewis 2008).
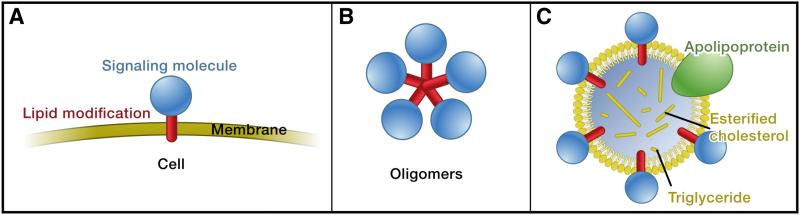
Signaling molecules (blue) are often modified by lipid attachments (red), and they can be inserted into membranes (A). In order to act on cells at a distance from the producing cell, these signaling molecules have to move through a hydrophilic environment. Formation of oligomers (B) and lipoprotein particles (C) are thought to mask hydrophobic residues or modifications and have been implicated in the transport of hydrophobic signals such as Hh and Wg. Figure modified from (Eaton 2008).
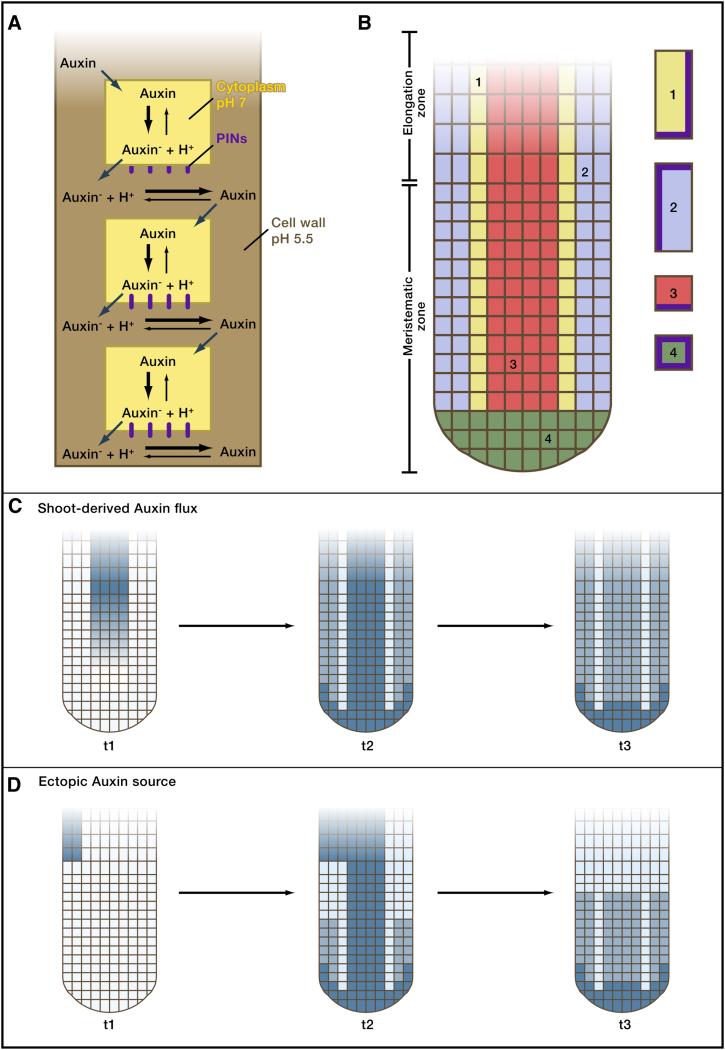
A) Polar auxin transport. Auxin can diffuse in the cell wall (brown) and enter cells. However, once inside the less acidic environment of the cells, Auxin becomes deprotonated (Auxin-) and can no longer leave the cell passively. PINs (red) are specific transport proteins that carry anionic Auxin- out of the cell. PINs are highly localized, often to the base of cells, and thereby lead to a directional transport of Auxin. Figure modified from (Robert and Friml 2009). B) Root architecture and localization of PINs. The Arabidopsis root consists of a meristematic zone, where growth occurs, and an elongation zone. The vasculature is indicated in red, epidermal layers in blue, border cells in yellow and columella tiers in cyan. All cells are surrounded by a cell wall (green). The localization of PIN proteins (brown) in cells within the indicated subregions is shown. Figure modified from (Grieneisen et al. 2007). C) Establishment and maintenance of an Auxin (blue) concentration maximum in a root with shoot-derived Auxin flux. Three time points of computational simulations are shown. Auxin flows through the phloem from the shoot to the root (t1). The distribution of PINs concentrates Auxin at the root tip (t2) and can maintain the Auxin concentration maximum even when the shoot-derived Auxin flux ceases (t3). Figure modified from (Grieneisen et al. 2007). D) Accumulation of Auxin (blue) at the root meristem from localized ectopic Auxin production. Three time points of computational simulations are shown. Localized Auxin production from a single cell at timepoint t1 is sufficient to generate an Auxin maximum at the root tip (t3) due to the distribution and subcellular localization of PINs. Figure modified from (Grieneisen et al. 2007).
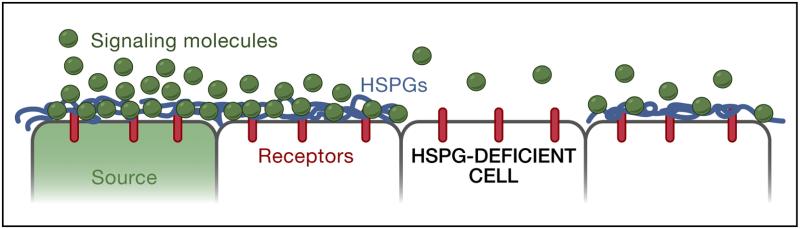
A) Interactions of signaling molecules with HSPGs. HSPGs are often associated with epithelial cell surfaces. Binding to HSPGs can alter the mobility of a signal, concentrate ligand at the surface of cells, promote or hinder ligand-receptor interactions, and influence the extracellular stability of a ligand. For example, in the absence of HSPGs, signaling molecules may not be retained on the cell surface and thereby fail to travel to the next cells. B) and C) Differential mobility of a signaling molecule in two different tissues. B) The embryonic wing (middle) and haltere discs (right) give rise to the adult wing and haltere, respectively (left). C) Dpp is more mobile in the embryonic wing disc than the haltere disc. The Dpp, Tkv, P-Mad (indicative of signaling range) and HSPG distributions in the wing disc (blue) and haltere disc (red) are shown. It is thought that Dpp movement in the haltere disc is restricted due to Ubx-mediated upregulation of the Dpp receptor Tkv in the medial domain of the disc. Ubx-mediated asymmetric expression of the HSPG core protein Dally is thought to bias Dpp diffusion anteriorly in the haltere disc. Note that Ubx also downregulates dpp transcription leading to decreased Dpp levels in the haltere disc (Crickmore and Mann 2006; de Navas et al. 2006; Crickmore and Mann 2007; Makhijani et al. 2007; Wartlick et al. 2011). Figure modified from (Crickmore and Mann 2008).
References
-
- Annes JP, Munger JS, Rifkin DB. Making sense of latent TGFβ activation. J Cell Sci. 2003;116:217–224. - PubMed
-
- Ayers KL, Gallet A, Staccini-Lavenant L, Therond PP. The long-range activity of Hedgehog is regulated in the apical extracellular space by the glypican Dally and the hydrolase Notum. Dev Cell. 2010;18:605–620. - PubMed
-
- Baeg GH, Lin X, Khare N, Baumgartner S, Perrimon N. Heparan sulfate proteoglycans are critical for the organization of the extracellular distribution of Wingless. Development. 2001;128:87–94. - PubMed
-
- Baeg GH, Selva EM, Goodman RM, Dasgupta R, Perrimon N. The Wingless morphogen gradient is established by the cooperative action of Frizzled and Heparan Sulfate Proteoglycan receptors. Dev Biol. 2004;276:89–100. - PubMed
-
- Bahlmann F, Wellek S, Reinhardt I, Krummenauer F, Merz E, Welter C. Reference values of fetal aortic flow velocity waveforms and associated intra-observer reliability in normal pregnancies. Ultrasound Obstet Gynecol. 2001;17:42–49. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
