

Minocycline attenuates photoreceptor degeneration in a mouse model of subretinal hemorrhage microglial: inhibition as a potential therapeutic strategy
- PMID: 21763674
- PMCID: PMC3157282
- DOI: 10.1016/j.ajpath.2011.05.042
Minocycline attenuates photoreceptor degeneration in a mouse model of subretinal hemorrhage microglial: inhibition as a potential therapeutic strategy
Abstract
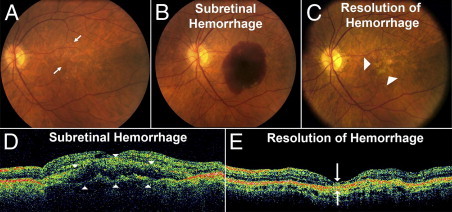
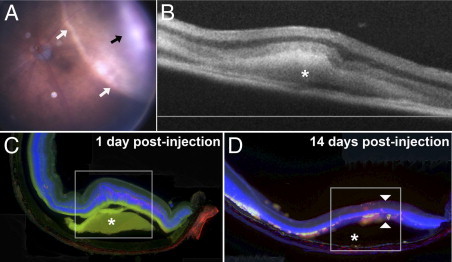
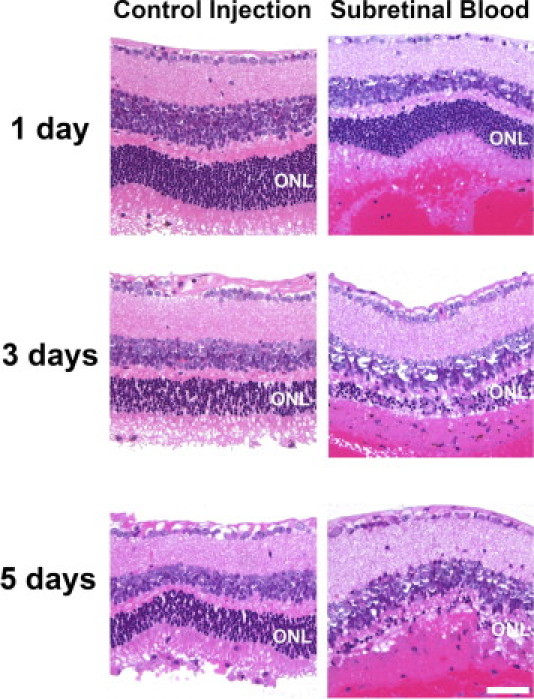
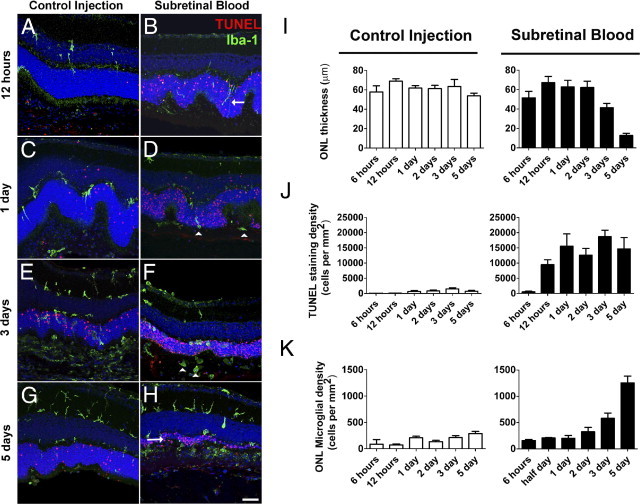
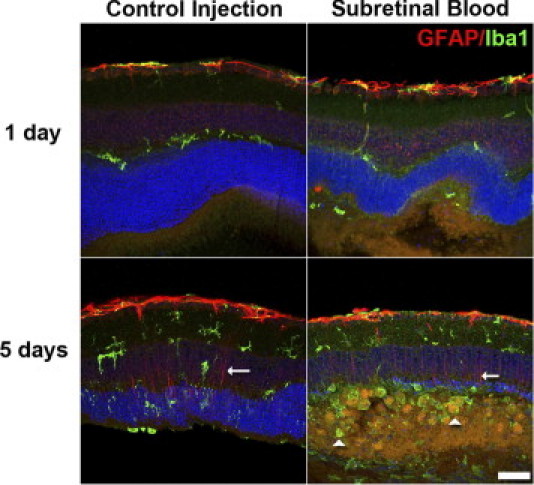
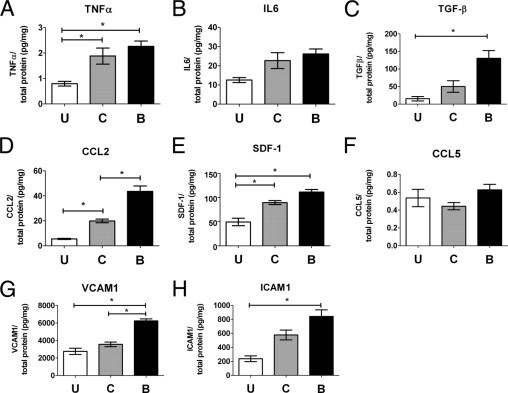
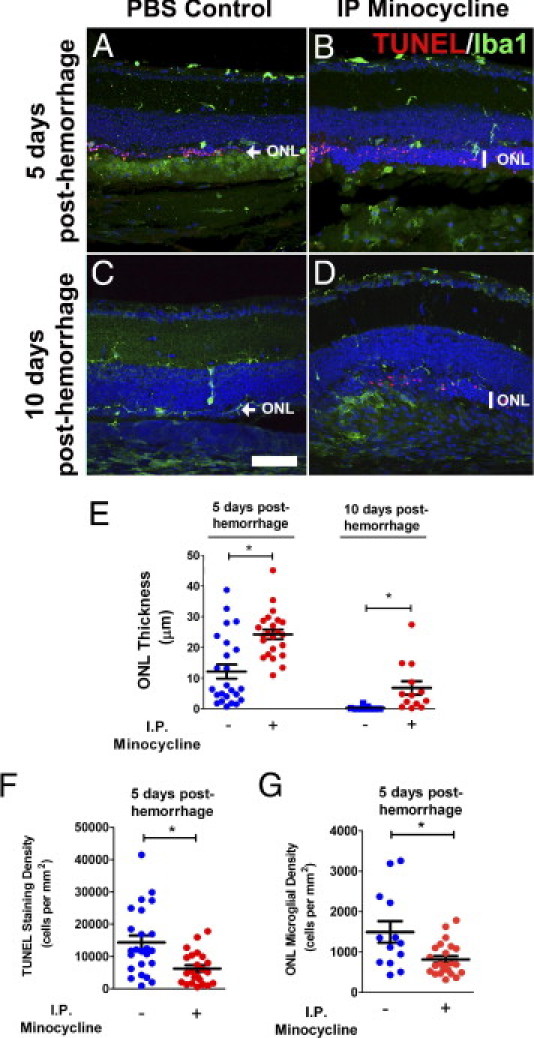
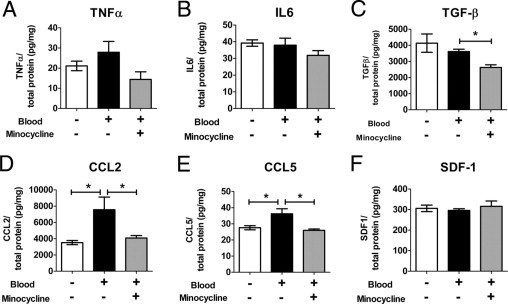
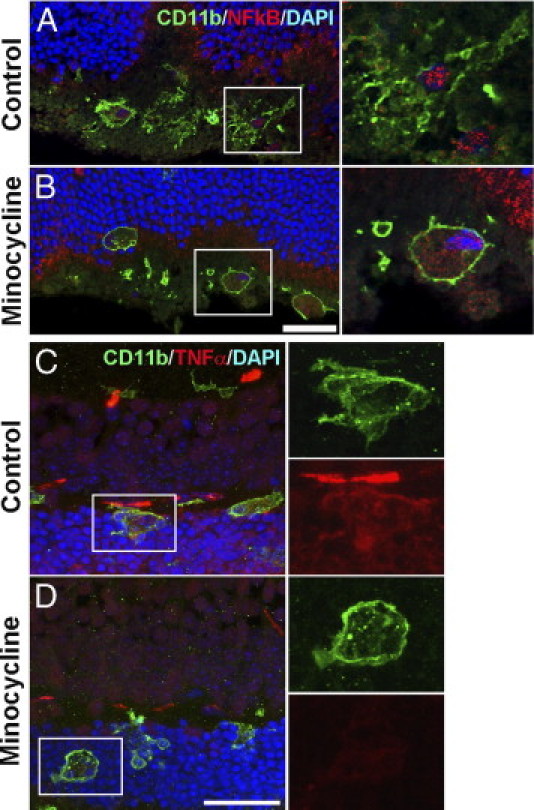
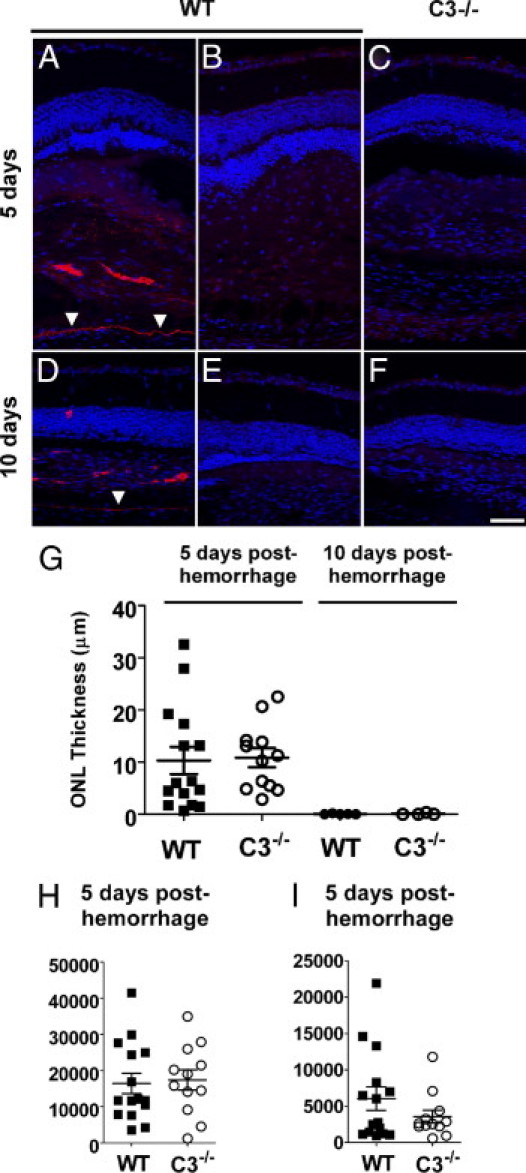
Hemorrhage under the neural retina (subretinal hemorrhage) can occur in the context of age-related macular degeneration and induce subsequent photoreceptor cell death and permanent vision loss. Current treatments with the objective of removing or displacing the hemorrhage are invasive and of mixed efficacy. We created a mouse model of subretinal hemorrhage to characterize the inflammatory responses and photoreceptor degeneration that occur in the acute aftermath of hemorrhage. It was observed that microglial infiltration into the outer retina commences as early as 6 hours after hemorrhage. Inflammatory cells progressively accumulate in the outer nuclear layer concurrently with photoreceptor degeneration and apoptosis. Administration of minocycline, an inhibitor of microglial activation, decreased microglial expression of chemotactic cytokines in vitro and reduced microglial infiltration and photoreceptor cell loss after subretinal hemorrhage in vivo. Inflammatory responses and photoreceptor atrophy occurred after subretinal hemorrhage, however, the degree of response and atrophy were similar between C3-deficient and C3-sufficient mice, indicating a limited role for complement-mediated processes. Our data indicate a role for inflammatory responses in driving photoreceptor cell loss in subretinal hemorrhage, and it is proposed that microglial inhibition may be beneficial in the treatment of subretinal hemorrhage.
Published by Elsevier Inc.
Figures
References
-
- Congdon N., O'Colmain B., Klaver C.C., Klein R., Munoz B., Friedman D.S., Kempen J., Taylor H.R., Mitchell P. Causes and prevalence of visual impairment among adults in the United States. Arch Ophthalmol. 2004;122:477–485. - PubMed
-
- Rein D.B., Wittenborn J.S., Zhang X., Honeycutt A.A., Lesesne S.B., Saaddine J. Forecasting age-related macular degeneration through the year 2050: the potential impact of new treatments. Arch Ophthalmol. 2009;127:533–540. - PubMed
-
- Kawasaki R., Yasuda M., Song S.J., Chen S.J., Jonas J.B., Wang J.J., Mitchell P., Wong T.Y. The prevalence of age-related macular degeneration in Asians: a systematic review and meta-analysis. Ophthalmology. 2010;117:921–927. - PubMed
-
- Mousa S.A., Mousa S.S. Current status of vascular endothelial growth factor inhibition in age-related macular degeneration. BioDrugs. 2010;24:183–194. - PubMed
-
- Bressler N.M. Antiangiogenic approaches to age-related macular degeneration today. Ophthalmology. 2009;116:S15–S23. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
