

Dynamic transcriptional events in embryonic stem cells mediated by the super elongation complex (SEC)
- PMID: 21764852
- PMCID: PMC3143939
- DOI: 10.1101/gad.2059211
Dynamic transcriptional events in embryonic stem cells mediated by the super elongation complex (SEC)
Abstract
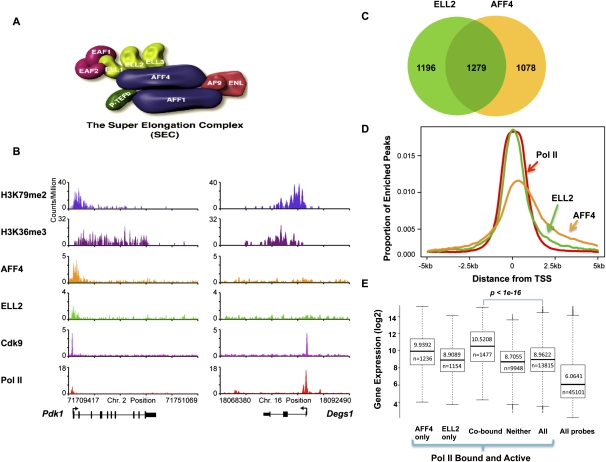
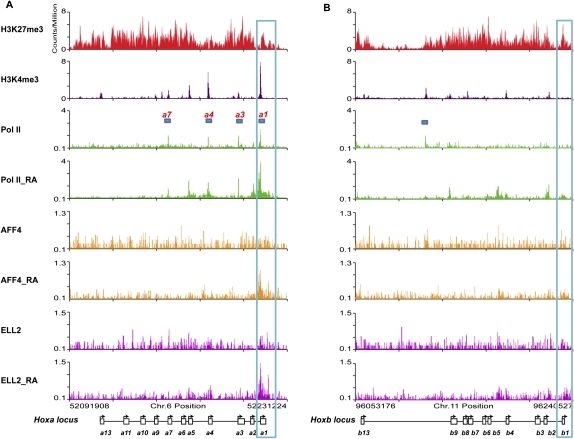
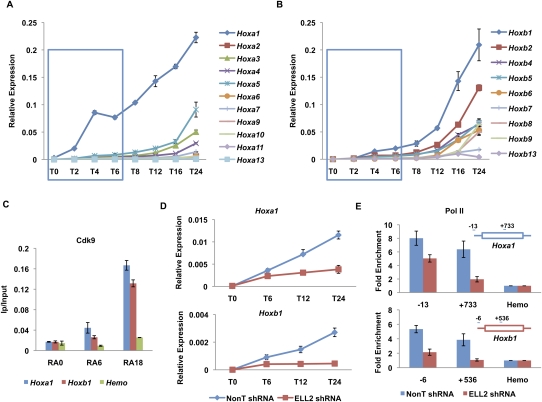
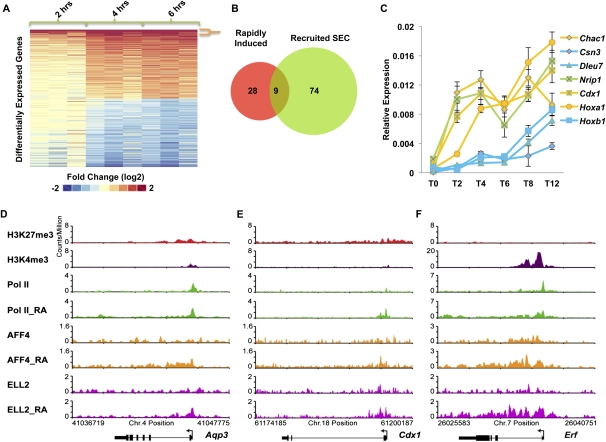
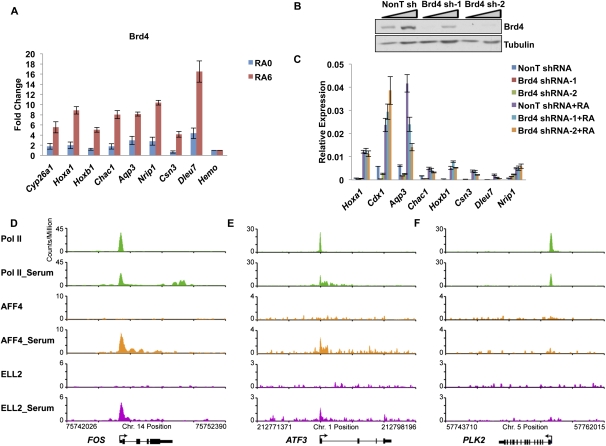
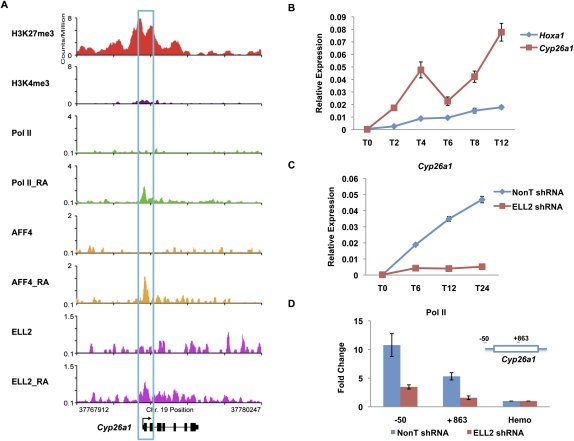
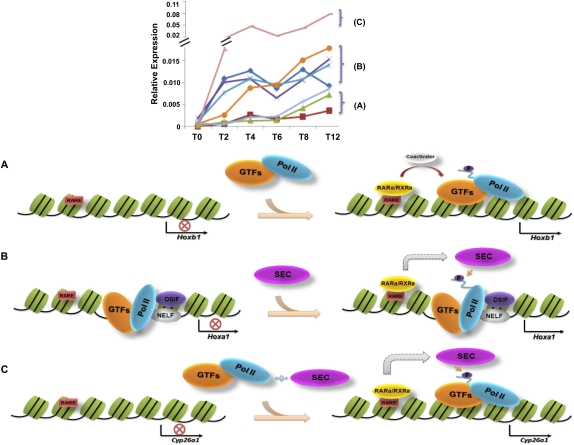
Transcriptional regulation of developmentally controlled genes is at the heart of differentiation and organogenesis. In this study, we performed global genomic analyses in murine embryonic stem (ES) cells and in human cells in response to activation signals. We identified an essential role for the ELL (eleven-nineteen lysine-rich leukemia gene)/P-TEFb (positive transcription elongation factor)-containing super elongation complex (SEC) in the regulation of gene expression, including several genes bearing paused RNA polymerase II (Pol II). Paused Pol II has been proposed to be associated with loci that respond rapidly to environmental stimuli. However, our studies in ES cells also identified a requirement for SEC at genes without paused Pol II, which also respond dynamically to differentiation signals. Our findings suggest that SEC is a major class of active P-TEFb-containing complexes required for transcriptional activation in response to environmental cues such as differentiation signals.
Figures
References
-
- Alexander T, Nolte C, Krumlauf R 2009. Hox genes and segmentation of the hindbrain and axial skeleton. Annu Rev Cell Dev Biol 25: 431–456 - PubMed
-
- Bernstein BE, Mikkelsen TS, Xie X, Kamal M, Huebert DJ, Cuff J, Fry B, Meissner A, Wernig M, Plath K, et al. 2006. A bivalent chromatin structure marks key developmental genes in embryonic stem cells. Cell 125: 315–326 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases