

Roles of the redox-active disulfide and histidine residues forming a catalytic dyad in reactions catalyzed by 2-ketopropyl coenzyme M oxidoreductase/carboxylase
- PMID: 21764916
- PMCID: PMC3165643
- DOI: 10.1128/JB.05231-11
Roles of the redox-active disulfide and histidine residues forming a catalytic dyad in reactions catalyzed by 2-ketopropyl coenzyme M oxidoreductase/carboxylase
Abstract
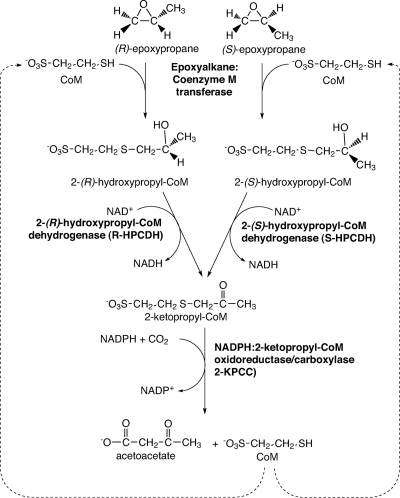
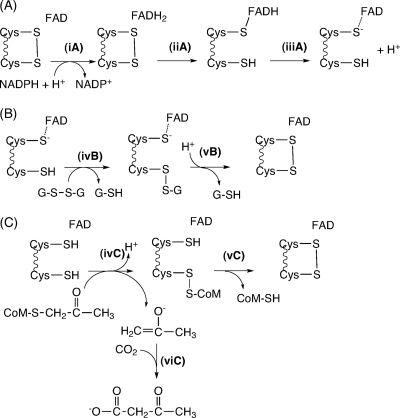
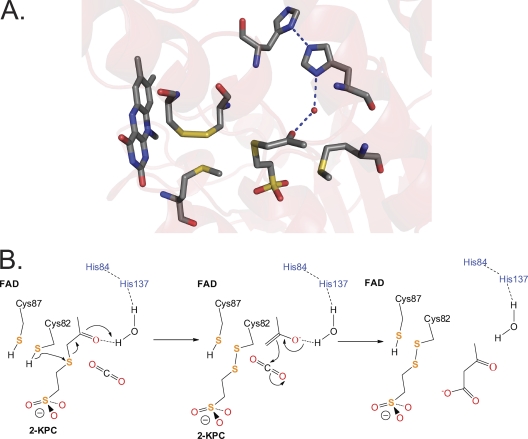
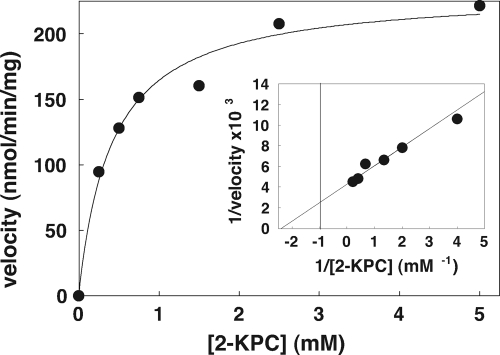
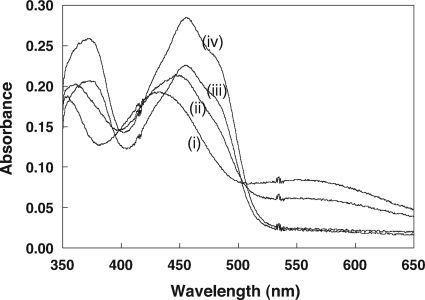
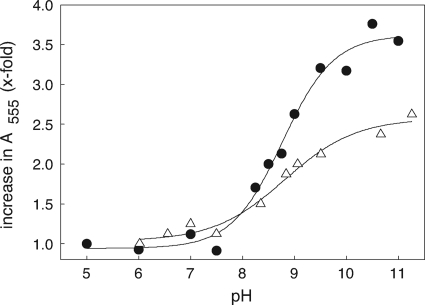
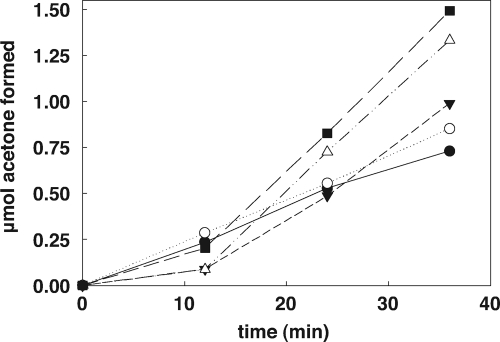
NADPH:2-ketopropyl-coenzyme M oxidoreductase/carboxylase (2-KPCC), an atypical member of the disulfide oxidoreductase (DSOR) family of enzymes, catalyzes the reductive cleavage and carboxylation of 2-ketopropyl-coenzyme M [2-(2-ketopropylthio)ethanesulfonate; 2-KPC] to form acetoacetate and coenzyme M (CoM) in the bacterial pathway of propylene metabolism. Structural studies of 2-KPCC from Xanthobacter autotrophicus strain Py2 have revealed a distinctive active-site architecture that includes a putative catalytic triad consisting of two histidine residues that are hydrogen bonded to an ordered water molecule proposed to stabilize enolacetone formed from dithiol-mediated 2-KPC thioether bond cleavage. Site-directed mutants of 2-KPCC were constructed to test the tenets of the mechanism proposed from studies of the native enzyme. Mutagenesis of the interchange thiol of 2-KPCC (C82A) abolished all redox-dependent reactions of 2-KPCC (2-KPC carboxylation or protonation). The air-oxidized C82A mutant, as well as wild-type 2-KPCC, exhibited the characteristic charge transfer absorbance seen in site-directed variants of other DSOR enzymes but with a pK(a) value for C87 (8.8) four units higher (i.e., four orders of magnitude less acidic) than that for the flavin thiol of canonical DSOR enzymes. The same higher pK(a) value was observed in native 2-KPCC when the interchange thiol was alkylated by the CoM analog 2-bromoethanesulfonate. Mutagenesis of the flavin thiol (C87A) also resulted in an inactive enzyme for steady-state redox-dependent reactions, but this variant catalyzed a single-turnover reaction producing a 0.8:1 ratio of product to enzyme. Mutagenesis of the histidine proximal to the ordered water (H137A) led to nearly complete loss of redox-dependent 2-KPCC reactions, while mutagenesis of the distal histidine (H84A) reduced these activities by 58 to 76%. A redox-independent reaction of 2-KPCC (acetoacetate decarboxylation) was not decreased for any of the aforementioned site-directed mutants. We interpreted and rationalized these results in terms of a mechanism of catalysis for 2-KPCC employing a unique hydrophobic active-site architecture promoting thioether bond cleavage and enolacetone formation not seen for other DSOR enzymes.
Copyright © 2011, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- Allen J. R., Ensign S. A. 1997. Purification to homogeneity and reconstitution of the individual components of the epoxide carboxylase multiprotein enzyme complex from Xanthobacter strain Py2. J. Biol. Chem. 272:32121–32128 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
