

Cytomegalovirus-specific T cell immunity is maintained in immunosenescent rhesus macaques
- PMID: 21765018
- PMCID: PMC3151292
- DOI: 10.4049/jimmunol.1100560
Cytomegalovirus-specific T cell immunity is maintained in immunosenescent rhesus macaques
Abstract
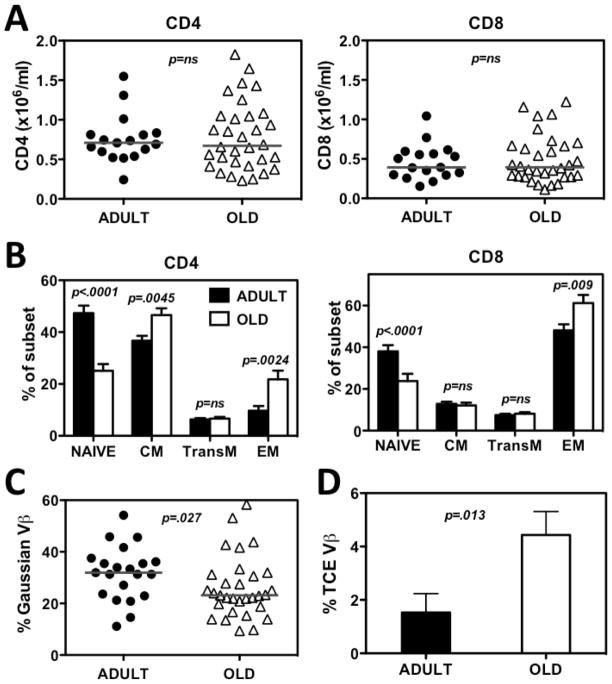
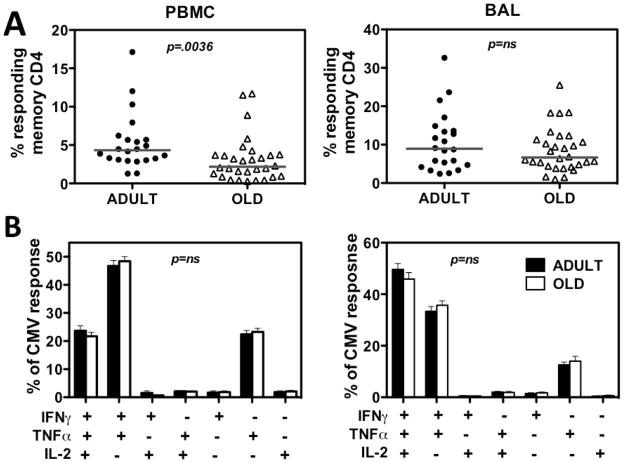
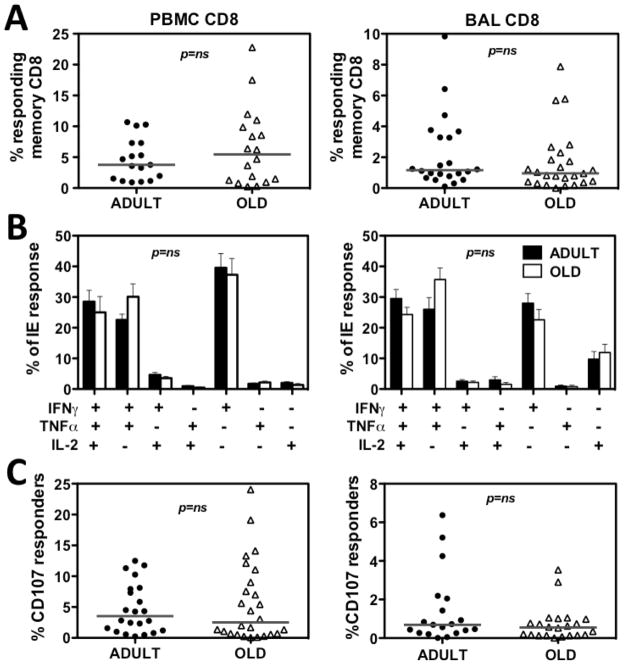
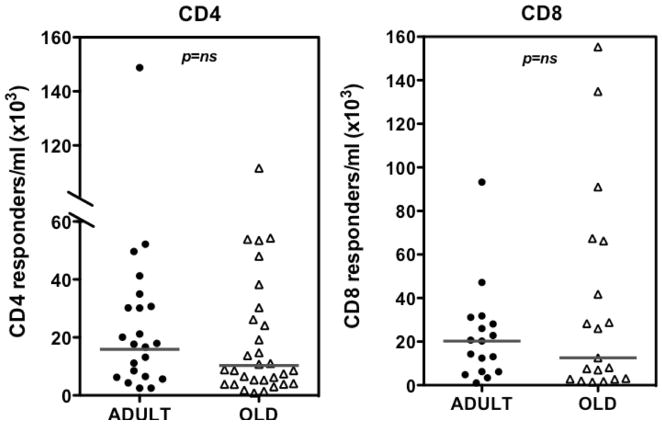
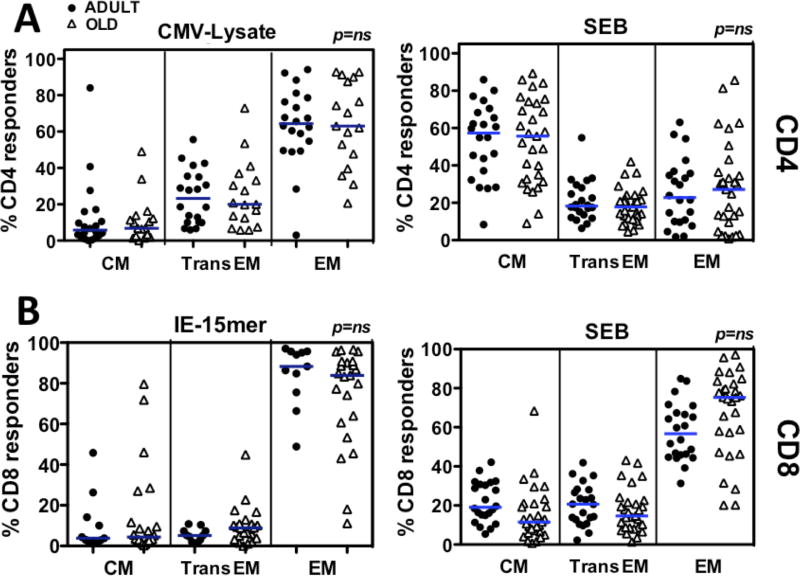
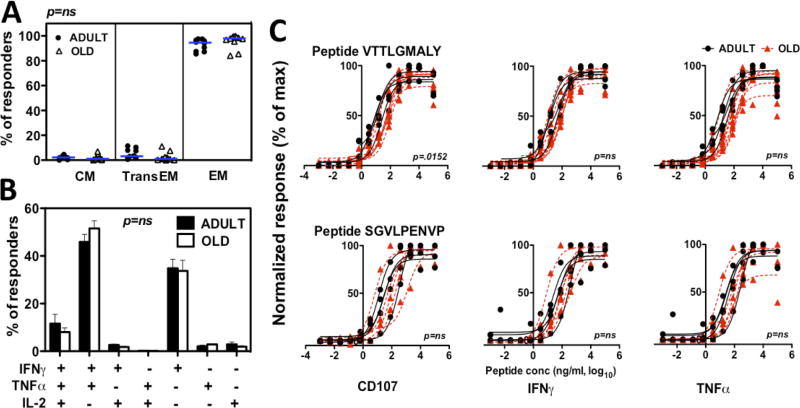
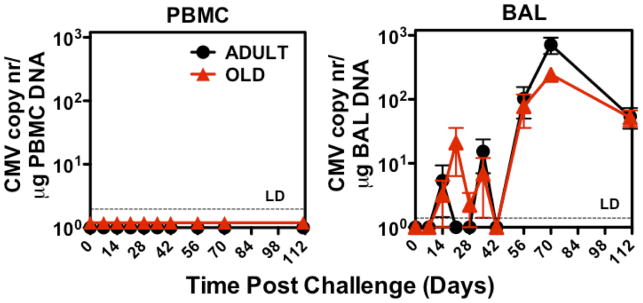
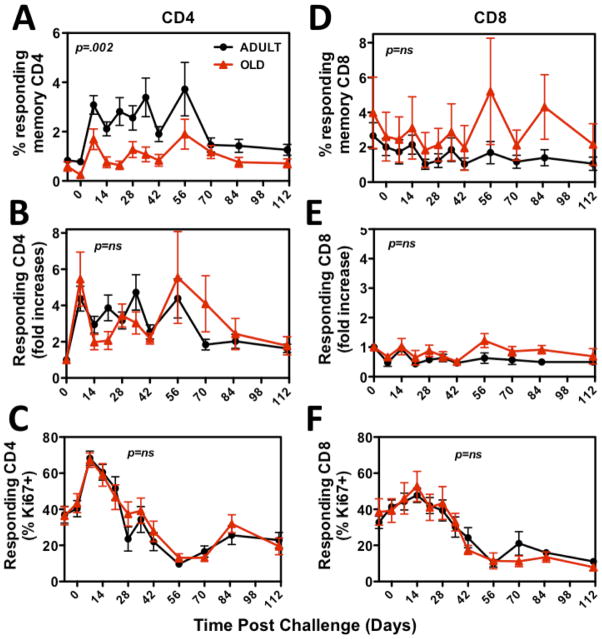
Although CMV infection is largely benign in immunocompetent people, the specific T cell responses associated with control of this persistent virus are enormous and must be maintained for life. These responses may increase with advanced age and have been linked to an "immune risk profile" that is associated with poor immune responsiveness and increased mortality in aged individuals. Based on this association, it has been suggested that CMV-specific T cell responses might become dysfunctional with age and thereby contribute to the development of immune senescence by homeostatic disruption of other T cell populations, diminished control of CMV replication, and/or excess chronic inflammation. In this study, we use the rhesus macaque (RM) model of aging to ask whether the quantity and quality of CMV-specific T cell responses differ between healthy adult RMs and elderly RMs that manifest hallmarks of immune aging. We demonstrate that the size of the CD4(+) and CD8(+) CMV-specific T cell pools are similar in adult versus old RMs and show essentially identical phenotypic and functional characteristics, including a dominant effector memory phenotype, identical patterns of IFN-γ, TNF-α, and IL-2 production and cytotoxic degranulation, and comparable functional avidities of optimal epitope-specific CD8(+) T cells. Most importantly, the response to and protection against an in vivo CMV challenge were identical in adult and aged RMs. These data indicate that CMV-specific T cell immunity is well maintained in old RMs and argue against a primary role for progressive dysfunction of these responses in the development of immune senescence.
Figures
References
-
- Ginaldi L, Loreto MF, Corsi MP, Modesti M, De Martinis M. Immunosenescence and infectious diseases. Microbes Infect. 2001;3:851–857. - PubMed
-
- High KP. Infection as a cause of age-related morbidity and mortality. Ageing Res Rev. 2004;3:1–14. - PubMed
-
- Hakim FT, Gress RE. Immunosenescence: deficits in adaptive immunity in the elderly. Tissue Antigens. 2007;70:179–189. - PubMed
-
- Weiskopf D, Weinberger B, Grubeck-Loebenstein B. The aging of the immune system. Transpl Int. 2009;22:1041–1050. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
