

Reengineering rate-limiting, millisecond enzyme motions by introduction of an unnatural amino acid
- PMID: 21767494
- PMCID: PMC3136797
- DOI: 10.1016/j.bpj.2011.05.039
Reengineering rate-limiting, millisecond enzyme motions by introduction of an unnatural amino acid
Abstract
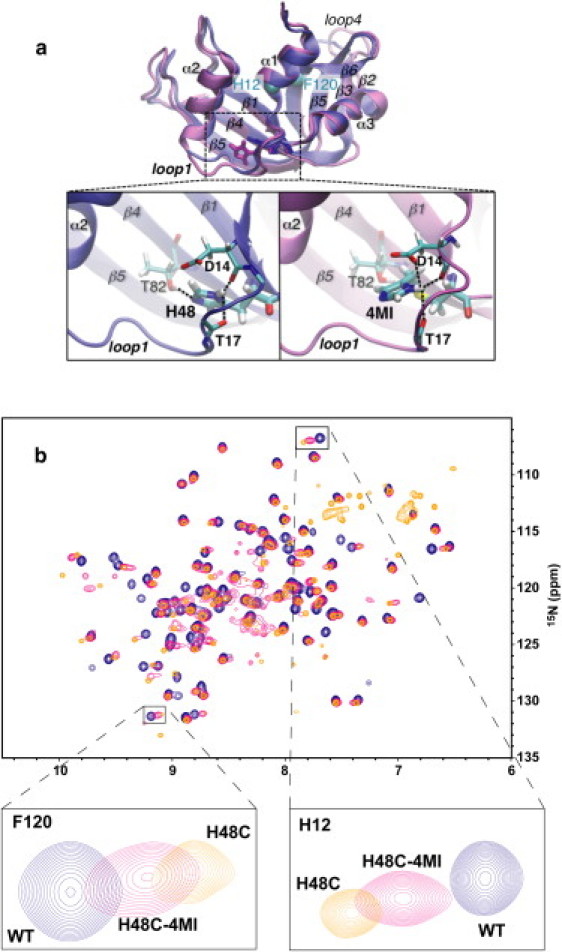
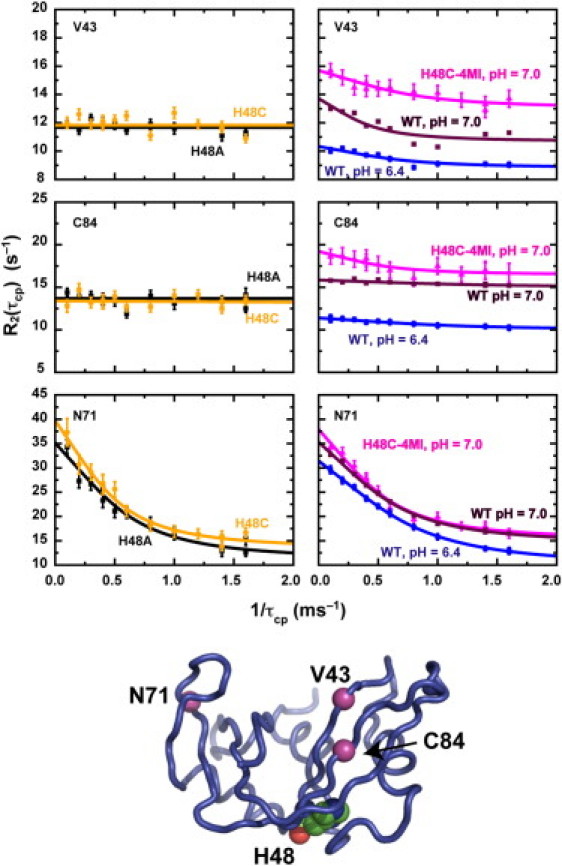
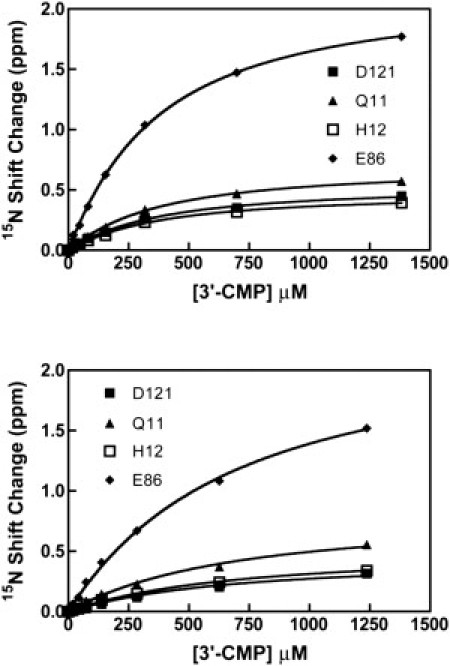
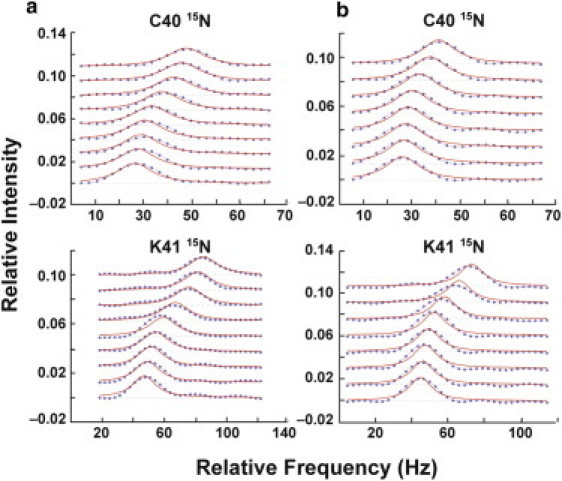
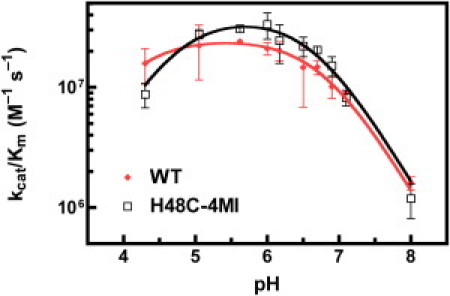
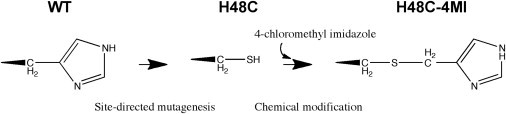
Rate-limiting millisecond motions in wild-type (WT) Ribonuclease A (RNase A) are modulated by histidine 48. Here, we incorporate an unnatural amino acid, thia-methylimidazole, at this site (H48C-4MI) to investigate the effects of a single residue on protein motions over multiple timescales and on enzyme catalytic turnover. Molecular dynamics simulations reveal that H48C-4MI retains some crucial WT-like hydrogen bonding interactions but the extent of protein-wide correlated motions in the nanosecond regime is decreased relative to WT. NMR Carr-Purcell-Meiboom-Gill relaxation dispersion experiments demonstrate that millisecond conformational motions in H48C-4MI are present over a similar pH range compared to WT. Furthermore, incorporation of this nonnatural amino acid allows retention of WT-like catalytic activity over the full pH range. These studies demonstrate that the complexity of the protein energy landscape during the catalytic cycle can be maintained using unnatural amino acids, which may prove useful in enzyme design efforts.
Copyright © 2011 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures

References
-
- Desjarlais J.R., Handel T.M. Side-chain and backbone flexibility in protein core design. J. Mol. Biol. 1999;290:305–318. - PubMed
-
- Harbury P.B., Plecs J.J., Kim P.S. High-resolution protein design with backbone freedom. Science. 1998;282:1462–1467. - PubMed
-
- Nauli S., Kuhlman B., Baker D. Computer-based redesign of a protein folding pathway. Nat. Struct. Biol. 2001;8:602–605. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
