

HSP70 mediates dissociation and reassociation of the 26S proteasome during adaptation to oxidative stress
- PMID: 21767633
- PMCID: PMC3172204
- DOI: 10.1016/j.freeradbiomed.2011.06.015
HSP70 mediates dissociation and reassociation of the 26S proteasome during adaptation to oxidative stress
Abstract
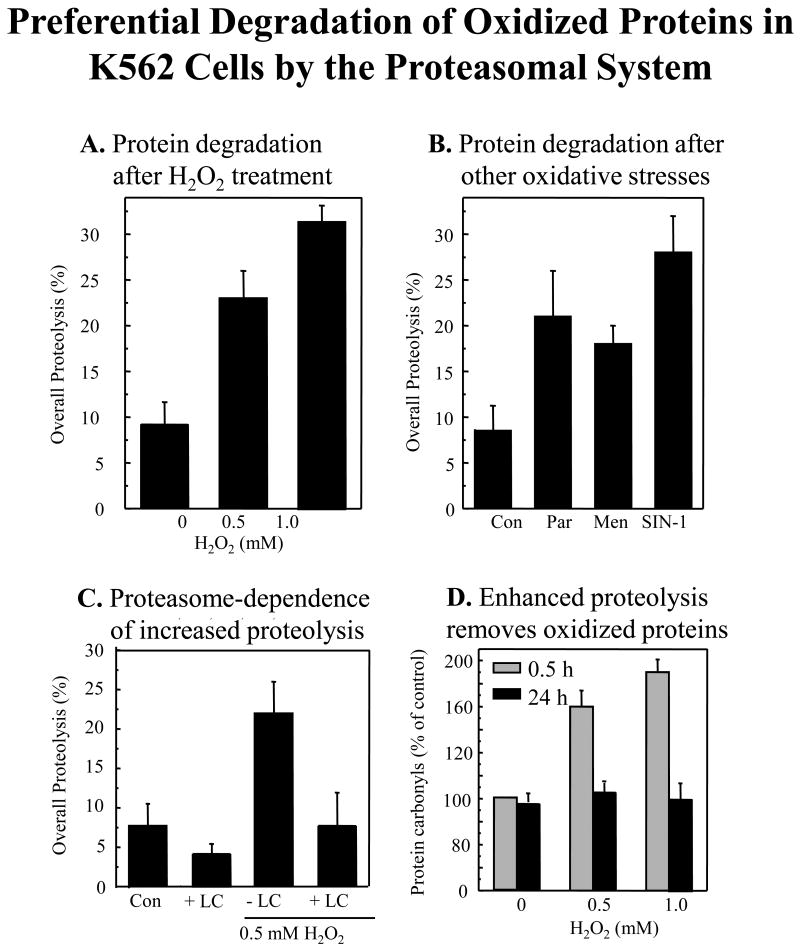
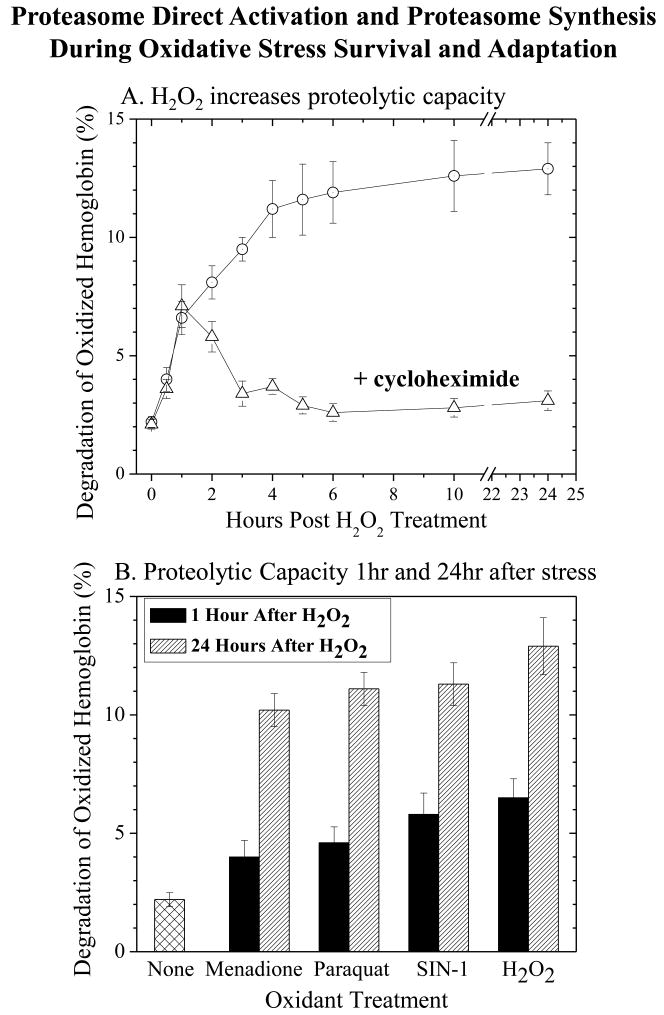
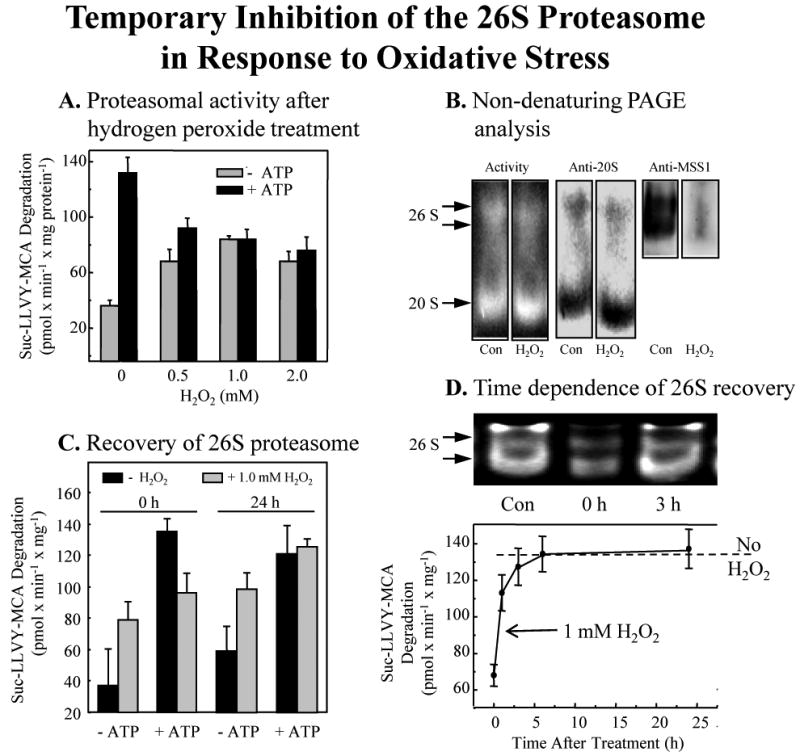
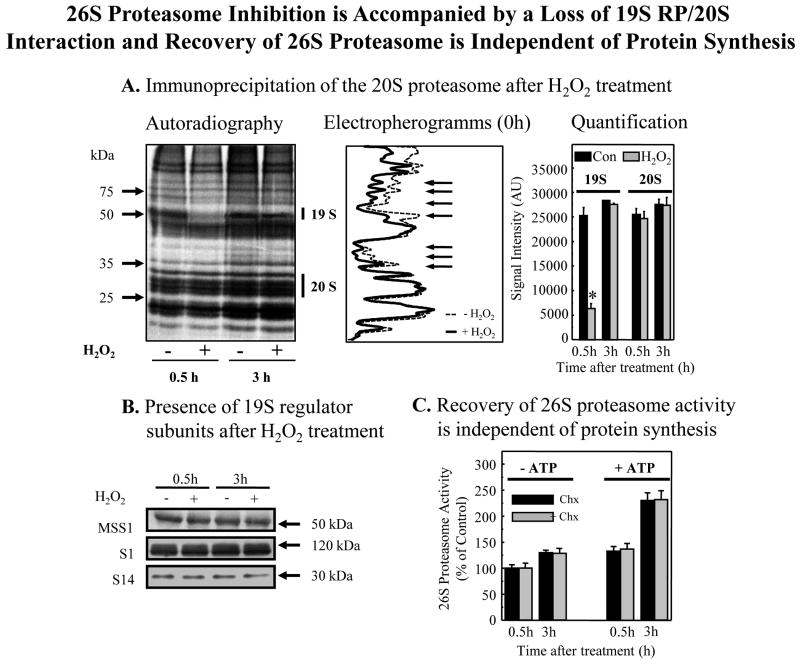
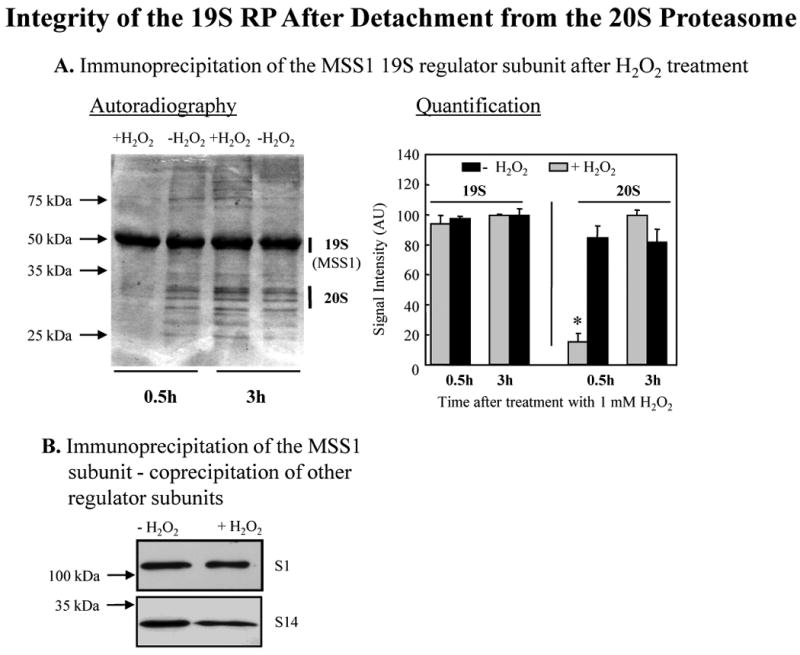
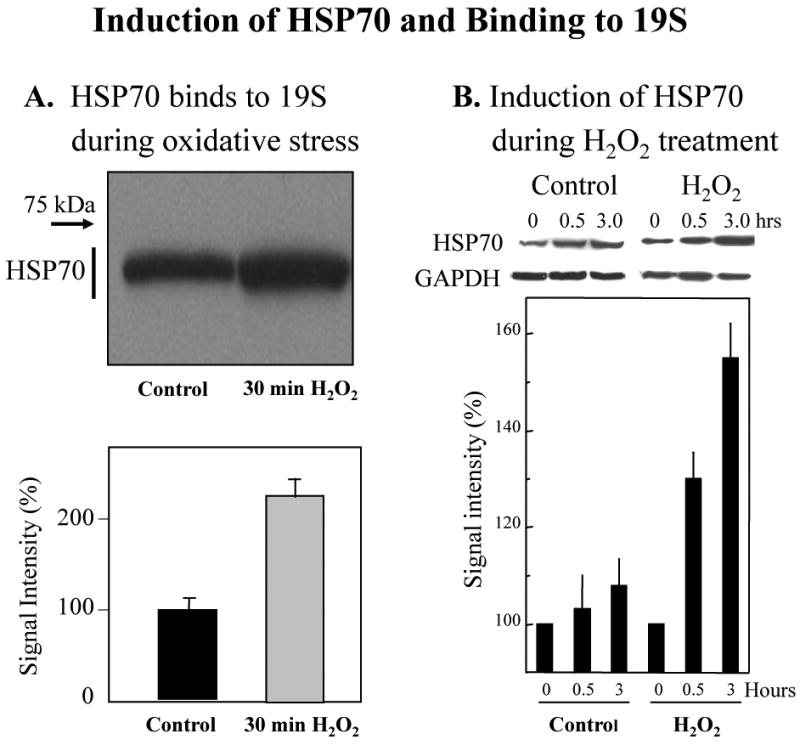
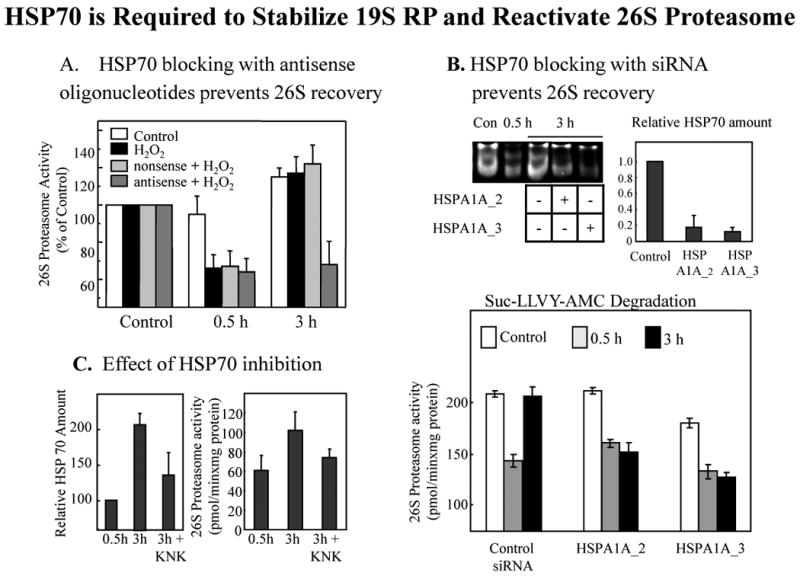
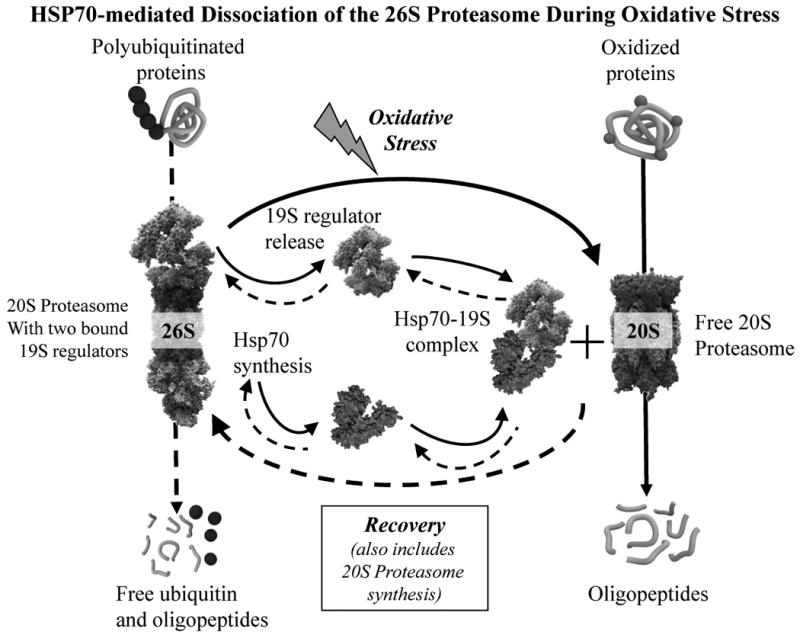
We report an entirely new role for the HSP70 chaperone in dissociating 26S proteasome complexes (into free 20S proteasomes and bound 19S regulators), preserving 19S regulators, and reconstituting 26S proteasomes in the first 1-3h after mild oxidative stress. These responses, coupled with direct 20S proteasome activation by poly(ADP ribose) polymerase in the nucleus and by PA28αβ in the cytoplasm, instantly provide cells with increased capacity to degrade oxidatively damaged proteins and to survive the initial effects of stress exposure. Subsequent adaptive (hormetic) processes (3-24h after stress exposure), mediated by several signal transduction pathways and involving increased transcription/translation of 20S proteasomes, immunoproteasomes, and PA28αβ, abrogate the need for 26S proteasome dissociation. During this adaptive period, HSP70 releases its bound 19S regulators, 26S proteasomes are reconstituted, and ATP-stimulated proteolysis is restored. The 26S proteasome-dependent, and ATP-stimulated, turnover of ubiquitinylated proteins is essential for normal cell metabolism, and its restoration is required for successful stress adaptation.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
References
-
- Tanaka K, Kasahara M. The MHC class I ligand-generating system: roles of immunoproteasomes and the interferon-gamma-inducible proteasome activator PA28. Immunol Rev. 1998;163:161–176. - PubMed
-
- Ma CP, Slaughter CA, DeMartino GN. Identification, purification, and characterization of a protein activator (PA28) of the 20 S proteasome (macropain) J Biol Chem. 1992;267:10515–10523. - PubMed
-
- Ahn K, Erlander M, Leturcq D, Peterson PA, Fruh K, Yang Y. In vivo characterization of the proteasome regulator PA28. J Biol Chem. 1996;271:18237–18242. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
