

Specific functions of lysosomal proteases in endocytic and autophagic pathways
- PMID: 21767668
- PMCID: PMC7105187
- DOI: 10.1016/j.bbapap.2011.07.003
Specific functions of lysosomal proteases in endocytic and autophagic pathways
Abstract
Endolysosomal vesicles form a highly dynamic multifunctional cellular compartment that contains multiple highly potent proteolytic enzymes. Originally these proteases have been assigned to cooperate solely in executing the unselective 'bulk proteolysis' within the acidic milieu of the lysosome. Although to some degree this notion still holds true, evidence is accumulating for specific and regulatory functions of individual 'acidic' proteases in many cellular processes linked to the endosomal/lysosomal compartment. Here we summarize and discuss the functions of individual endolysosomal proteases in such diverse processes as the termination of growth factor signaling, lipoprotein particle degradation, infection, antigen presentation, and autophagy. This article is part of a Special Issue entitled: Proteolysis 50 years after the discovery of lysosome.
Copyright © 2011 Elsevier B.V. All rights reserved.
Figures

 Active endolysosomal protease,
Active endolysosomal protease,  deficient endolysosomal protease,
deficient endolysosomal protease,  dimerized EGF receptors,
dimerized EGF receptors,  two EGF molecules.
two EGF molecules.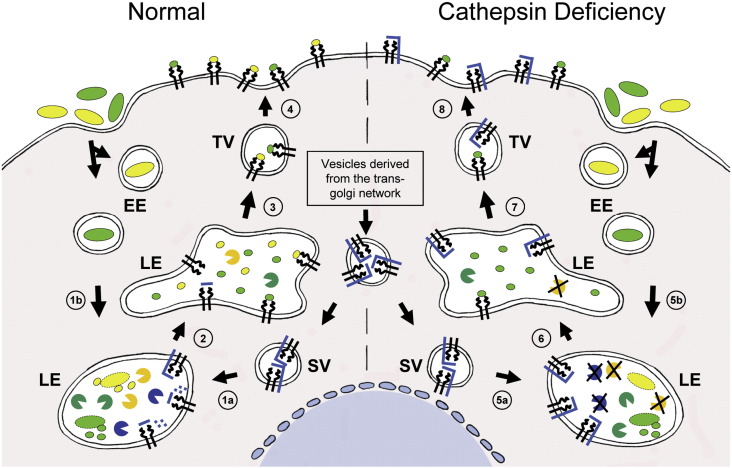 Active endolysosomal protease, deficient endolysosomal protease,
Active endolysosomal protease, deficient endolysosomal protease,  MHC class II molecule,
MHC class II molecule,  invariant chain,
invariant chain,  antigenic peptide,
antigenic peptide,  bacterium/pathogen/antigenic protein.
bacterium/pathogen/antigenic protein.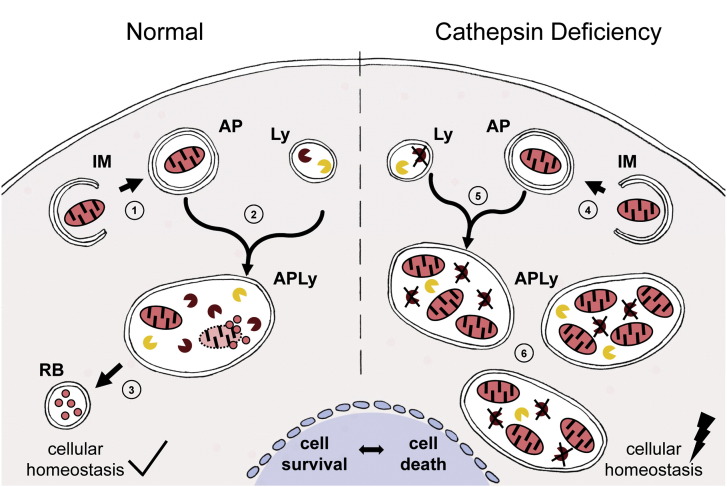 Active endolysosomal protease, deficient endolysosomal protease,
Active endolysosomal protease, deficient endolysosomal protease,  mitochondrion.
mitochondrion.References
-
- De Duve C., Wattiaux R. Functions of lysosomes. Annu. Rev. Physiol. 1966;28:435–492. - PubMed
-
- Willstätter R., Bamann E. Über die Proteasen der Magenschleimhaut. Erste Abhandlung über die Enzyme der Leukozyten., Hoppe-Seyler's. Z. Physiol. Chem. 1929;180:127–143.
-
- Goulet B., Baruch A., Moon N.S., Poirier M., Sansregret L.L., Erickson A., Bogyo M., Nepveu A. A cathepsin L isoform that is devoid of a signal peptide localizes to the nucleus in S phase and processes the CDP/Cux transcription factor. Mol. Cell. 2004;14:207–219. - PubMed
-
- Sever S., Altintas M.M., Nankoe S.R., Moller C.C., Ko D., Wei C., Henderson J., del Re E.C., Hsing L., Erickson A., Cohen C.D., Kretzler M., Kerjaschki D., Rudensky A., Nikolic B., Reiser J. Proteolytic processing of dynamin by cytoplasmic cathepsin L is a mechanism for proteinuric kidney disease. J. Clin. Invest. 2007;117:2095–2104. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
