

APC/CCdh1-dependent proteolysis of USP1 regulates the response to UV-mediated DNA damage
- PMID: 21768287
- PMCID: PMC3144416
- DOI: 10.1083/jcb.201101062
APC/CCdh1-dependent proteolysis of USP1 regulates the response to UV-mediated DNA damage
Abstract
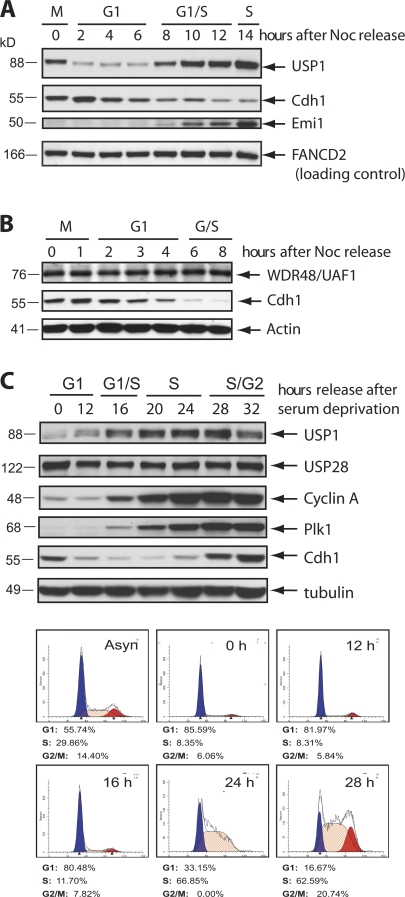
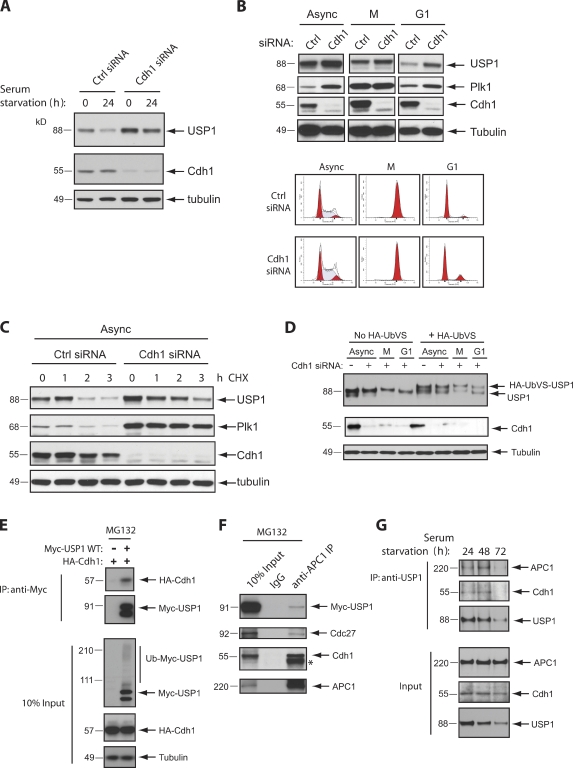
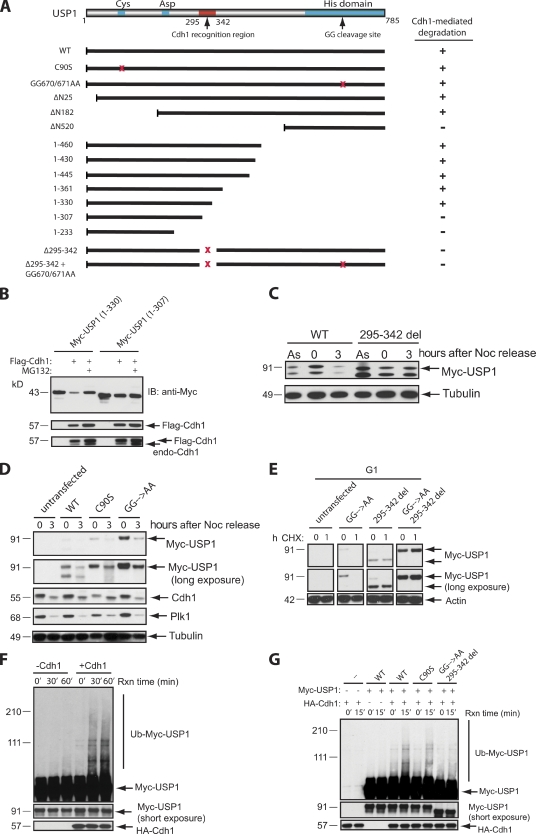
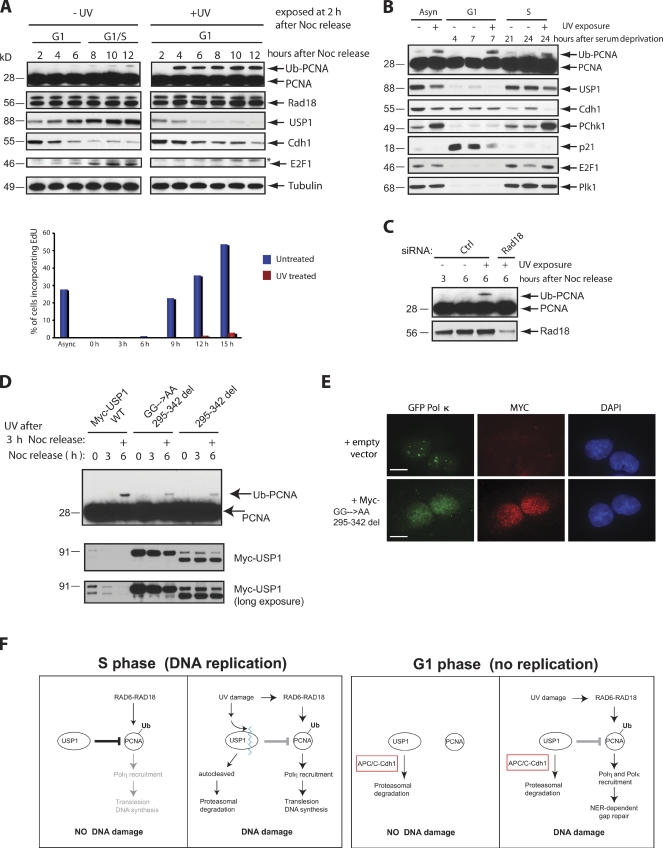
Targeted protein destruction of critical cellular regulators during the G1 phase of the cell cycle is achieved by anaphase-promoting complex/cyclosome(Cdh1) (APC/C(Cdh1)), a multisubunit E3 ubiquitin ligase. Cells lacking Cdh1 have been shown to accumulate deoxyribonucleic acid (DNA) damage, suggesting that it may play a previously unrecognized role in maintaining genomic stability. The ubiquitin-specific protease 1 (USP1) is a known critical regulator of DNA repair and genomic stability. In this paper, we report that USP1 was degraded in G1 via APC/C(Cdh1). USP1 levels were kept low in G1 to provide a permissive condition for inducing proliferating cell nuclear antigen (PCNA) monoubiquitination in response to ultraviolet (UV) damage before DNA replication. Importantly, expression of a USP1 mutant that cannot be degraded via APC/C(Cdh1) inhibited PCNA monoubiquitination during G1, likely compromising the recruitment of trans-lesion synthesis polymerase to UV repair sites. Thus, we propose a role for APC/C(Cdh1) in modulating the status of PCNA monoubiquitination and UV DNA repair before S phase entry.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
