

doi: 10.1182/blood-2011-04-348748.
Epub 2011 Jul 18.
A reliable ex vivo invasion assay of human reticulocytes by Plasmodium vivax
Affiliations
- PMID: 21768300
- PMCID: PMC3438884
- DOI: 10.1182/blood-2011-04-348748
Item in Clipboard
A reliable ex vivo invasion assay of human reticulocytes by Plasmodium vivax
Blood.
.
Abstract
Currently, there are no reliable RBC invasion assays to guide the discovery of vaccines against Plasmodium vivax, the most prevalent malaria parasite in Asia and South America. Here we describe a protocol for an ex vivo P vivax invasion assay that can be easily deployed in laboratories located in endemic countries. The assay is based on mixing enriched cord blood reticulocytes with matured, trypsin-treated P vivax schizonts concentrated from clinical isolates. The reliability of this assay was demonstrated using a large panel of P vivax isolates freshly collected from patients in Thailand.
Figures
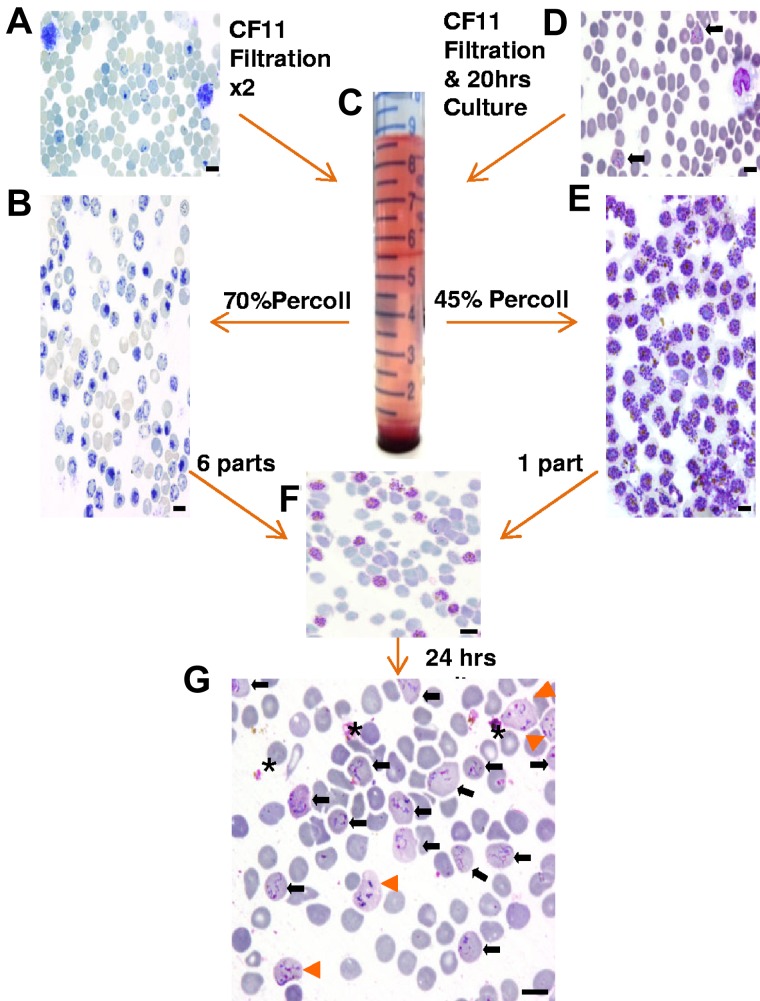
An overview of the assay methodology. Thin smears stained with new methylene blue (A-B) or Giemsa (D-G) illustrating the key methodologic steps of the ex vivo P vivax invasion assay. (A) Cord blood at collection contains a mixture of normocytes, reticulocytes, and leukocytes. (B) After 2 rounds of leukocyte depletion on a CF-11 cellulose column, reticulocytes form the majority of the cells present in the band (seen at the 5.5-mL mark on the tube pictured in panel C) obtained by enrichment on a 70% Percoll cushion. (D) Thin smear showing mature trophozoites (black arrows) and leukocytes typically observed from P vivax isolates collected from patients. (E) Typical yield of concentrated mature P vivax parasites obtained on a 45% Percoll cushion (from a band similar to the one depicted in panel C after a single round of leukocyte depletion on CF-11 and 20 hours ex vivo maturation). (F) The concentrated reticulocyte target cells (B) and mature P vivax schizonts (E) are mixed at a ratio of 6:1 for the invasion assay. (G) After culturing for ∼ 24 hours, the invasion assay mixture shows remnants of ruptured schizonts (*), and RBCs newly invaded by one (black arrow) or multiple merozoites (orange arrowhead). Black scale bar on the micrographs corresponds to 10 μm.
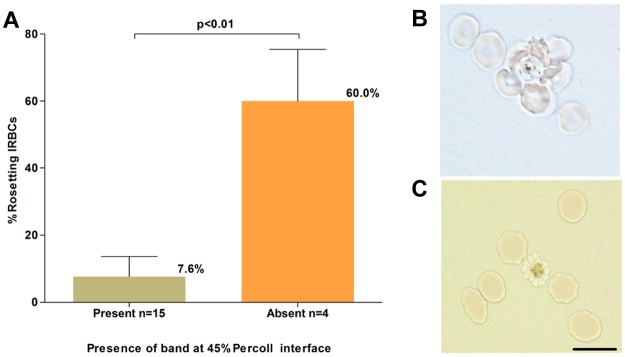
P vivax rosetting and its effect on schizont concentration. (A). The relationship between the percentage of rosetting P vivax IRBCs (in the 20-hour matured isolates) before concentration and presence or absence of a concentrated schizont band at the 45% Percoll interface. (B) A micrograph of an unstained wet preparation showing a P vivax rosette (more than one RBC attached to a central IRBC) collected from the pellet of a isolate that did not yield a band on the Percoll cushion. (C) Rosetting forms were not observed following treatment of the pellet with trypsin, and a band was subsequently obtained on the Percoll cushion. Black scale bar corresponds to 10 μm for both micrographs.
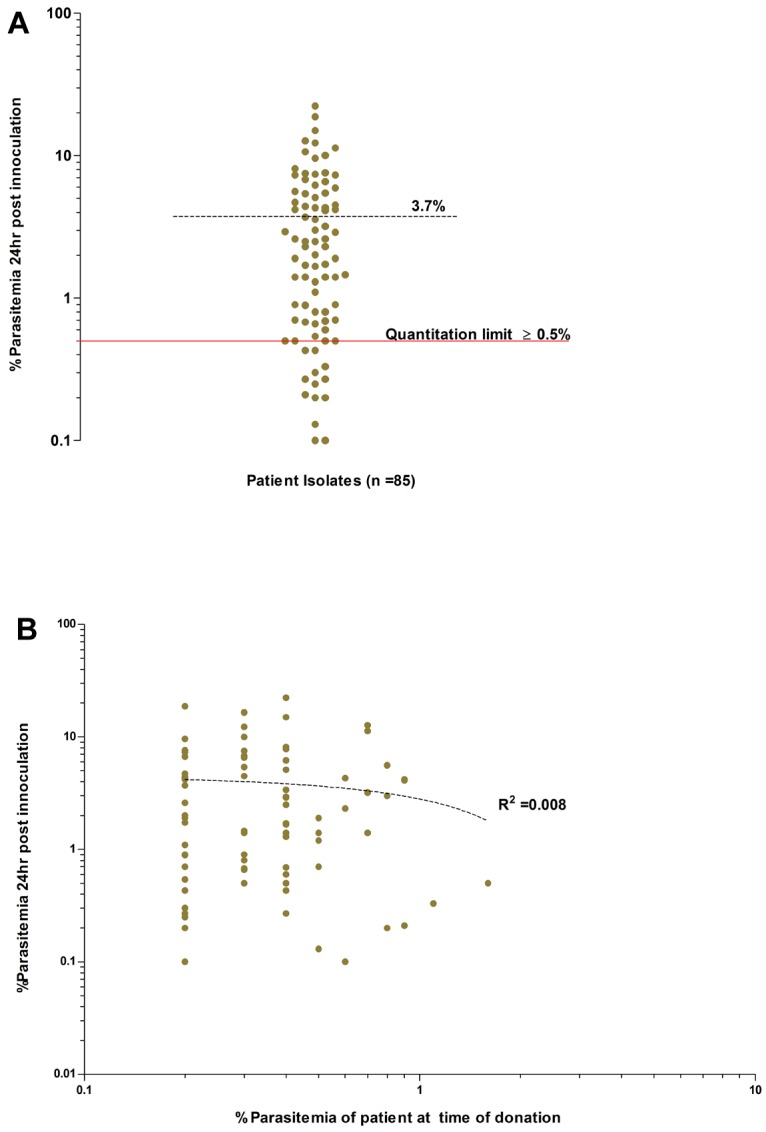
Overall invasion efficiencies of the P vivax field isolates used in the invasion assays. (A) Average invasion efficiency of enriched ex vivo–matured P vivax schizonts isolated from 85 isolates in concentrated reticulocytes collected from 28 cord blood isolates (experiments conducted in triplicate). The mean percentage of newly invaded RBCs (dashed black line) after ∼ 24 hours of incubation was 3.7% (95% CI: 2.8-4.7%, range 0.1-22.3%). Isolates with an invasion efficiency above the quantitation limit of ≥ 0.5% parasitemia (solid red line), provide those conducting assay with a 90% (power = 0.9) chance of detecting significance changes in parasitemia between 2 paired treatment groups (for example, Ab vs control). (B) Absence of any relationship between the ex vivo invasion efficiency and the in vivo admission parasitemia for each isolate (dotted line, goodness of fit: R2 = 0.008, P = .397).
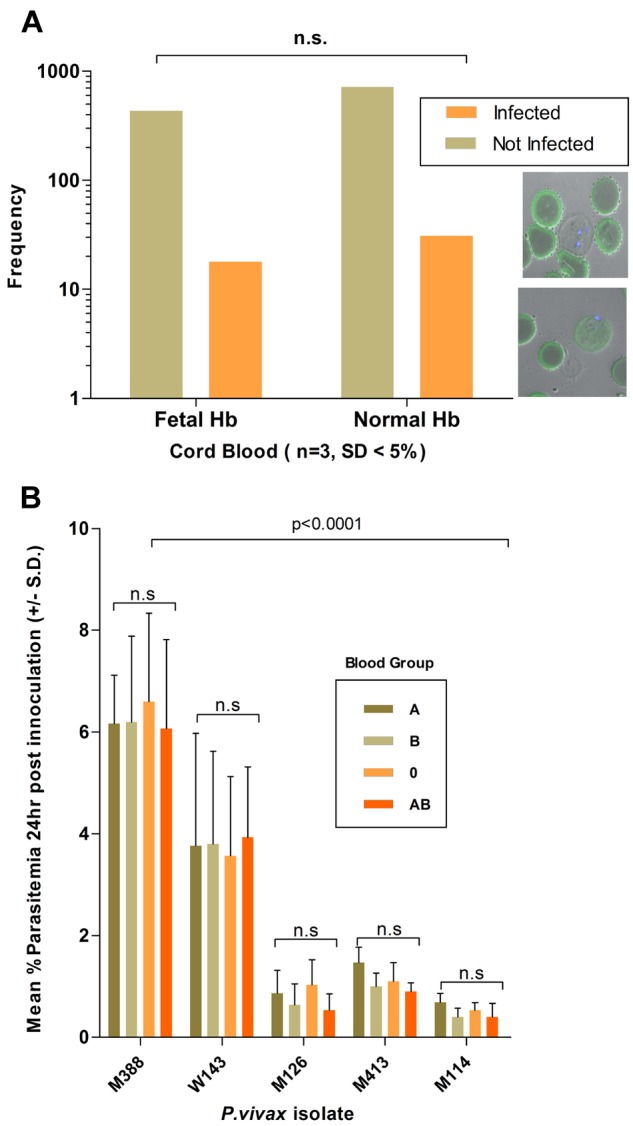
Effect of host cell attributes on invasion efficiency. (A) P vivax invasion efficiency, determined from 1200 target cells (n = 3 isolates), was not significantly associated with the presence of fetal Hb (stained in green). The bottom right inset shows a P vivax (nucleus stained with Dapi blue) in a newly invaded RBCs devoid (top micrograph) or containing fetal Hb. (B) The P vivax invasion efficiency (n = 5 isolates) was not affected by the ABO blood group of the reticulocytes used in the invasion assay. Invasion efficiencies were significantly associated with the particular isolate used (isolates M388 and W143 consistently invaded the distinct target cell preparation at a higher efficiency than that observed for isolates M126, M413, and M114).
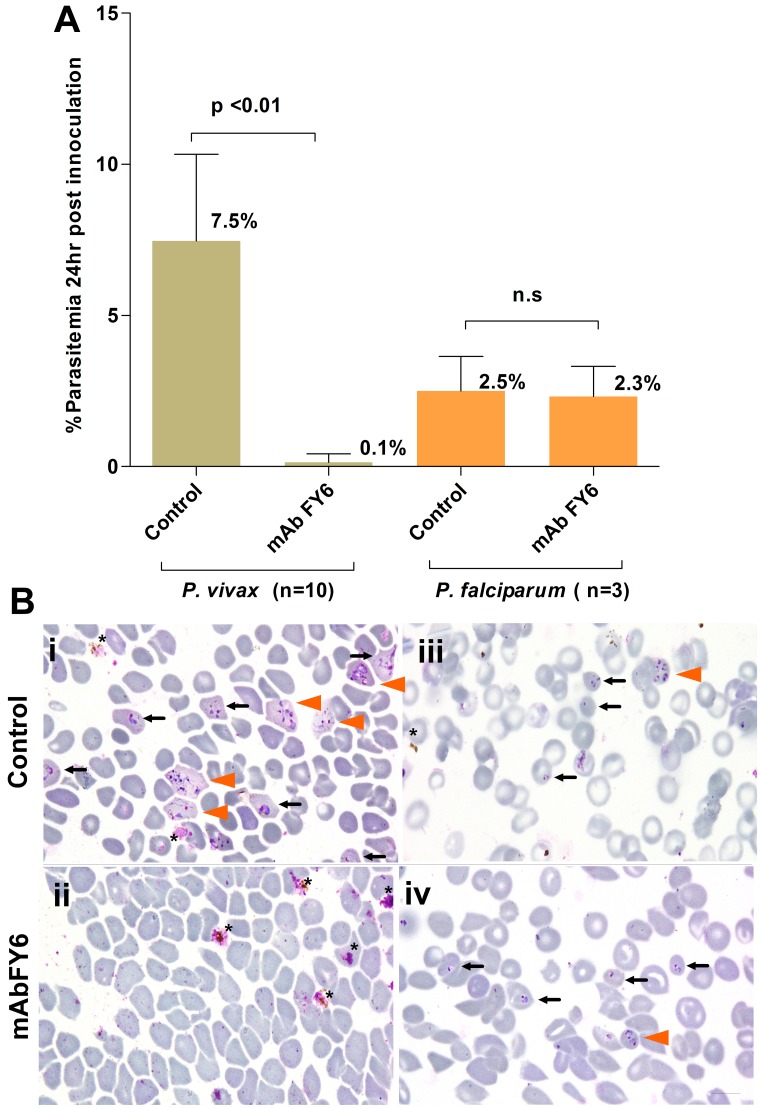
Inhibition of P vivax invasion by a mAb (25 μg/mL) specific to the 2C3 epitope of DARC, the obligatory receptor for P vivax merozoites. (A) Inhibition of P vivax (n = 10 isolates) invasion was near-total (mean 7.5% SD ± 2.9 vs 0.15% SD ± 0.28, P < .01, Wilcoxon sum of signed ranks [W] = 55.), whereas little inhibition was observed (P = .25, W = 6.0) when P falciparum (n = 3 isolates) were used in the invasion assay. The assays were conducted in triplicate for each isolate. (B) Representative micrographs are shown for the (i-ii) P vivax or (iii-iv) P falciparum invasion assays (annotations as in Figure 1). Remnants of ruptured schizonts (*), and RBCs newly invaded by 1 (black arrow) or multiple merozoites (orange arrowhead). Black scale bars on micrograph represent 10 μm.
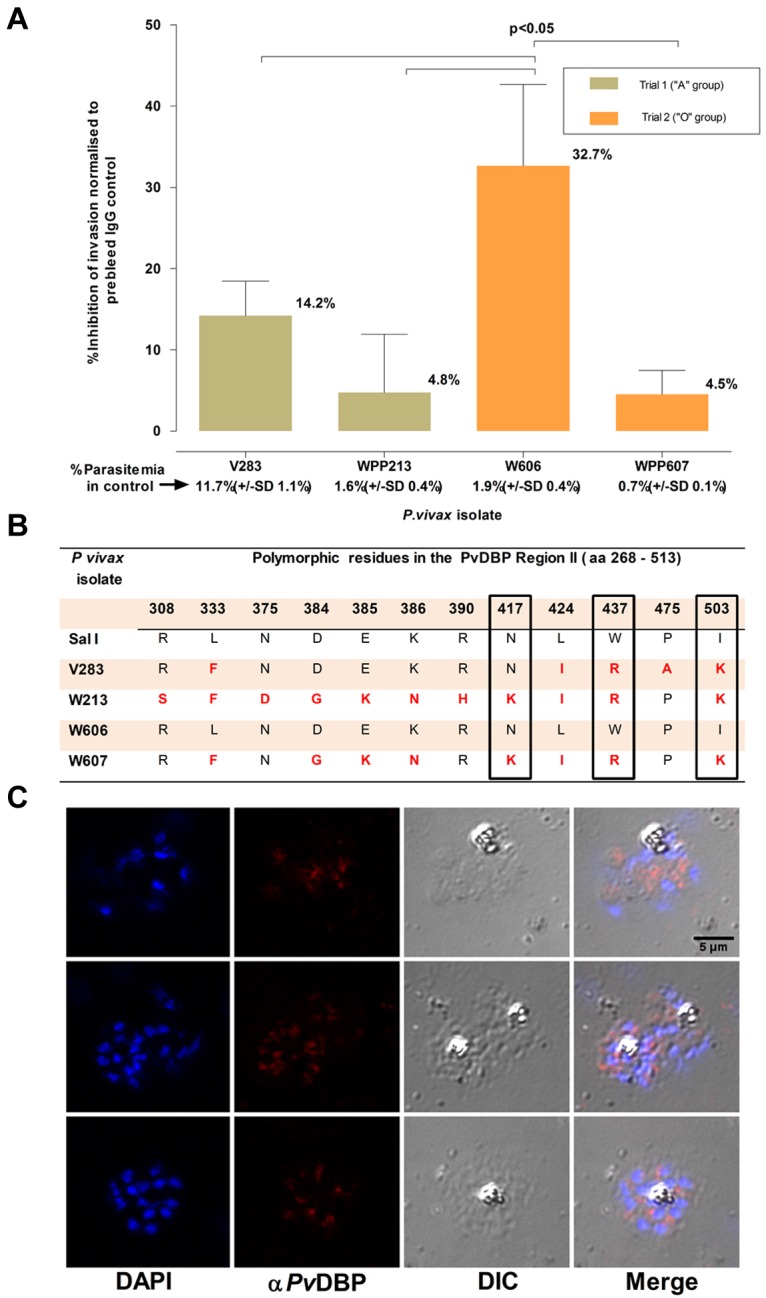
Inhibition of P vivax invasion by anti-PvDBP II IgG Abs. (A) The mean inhibition (percentage ± % SD) of anti–PvDBP II IgG Abs (100 μg/mL) on the P vivax invasion efficiency. The inhibition was normalized to invasion efficiencies obtained in the presence of IgG Abs (100 μg/mL) purified from a rabbit prebleed serum. The actual mean invasion efficiencies (percentage ± % SD) in the prebleed controls are included below the corresponding isolate identifier. The assays were conducted in triplicate on 4 isolates in 2 separate experiments involving reticulocyte concentrates from 2 cord blood samples of different ABO type. (B) Polymorphisms in the predicted amino acid sequence of the PvDBP region II region in the parasites present in the 4 isolates used in the assay. The sequences were compared with that present in the P vivax reference strain (Sal 1). Nonsynonymous mutations at 3 residues (417, 437, and 503) that were previously suggested to interfere with the inhibition of PvDBP region II binding to DARC are enclosed in black frames. (C) Immunofluorescence microscopy with anti–P vivax DBPII IgG reacted with merozoites from a P vivax isolate (3 examples given) used in this assay.
Comment in
-
"Baby" red cells to the rescue.Blood. 2011 Sep 29;118(13):3454-5. doi: 10.1182/blood-2011-08-370395. Blood. 2011. PMID: 21960678 No abstract available.
References
-
- Baird JK. Neglect of Plasmodium vivax malaria. Trends Parasitol. 2007;23(11):533–539. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
