

Reluctance to membrane binding enables accessibility of the synaptobrevin SNARE motif for SNARE complex formation
- PMID: 21768342
- PMCID: PMC3150886
- DOI: 10.1073/pnas.1105128108
Reluctance to membrane binding enables accessibility of the synaptobrevin SNARE motif for SNARE complex formation
Abstract
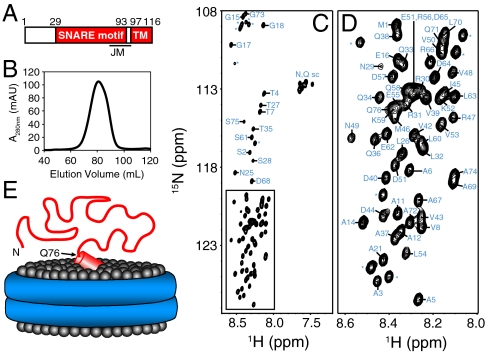
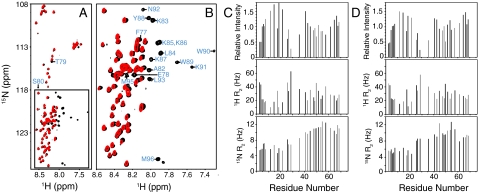
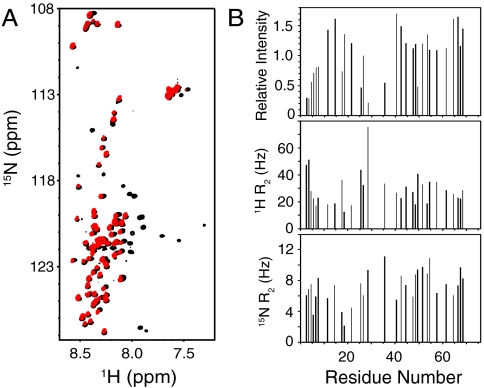
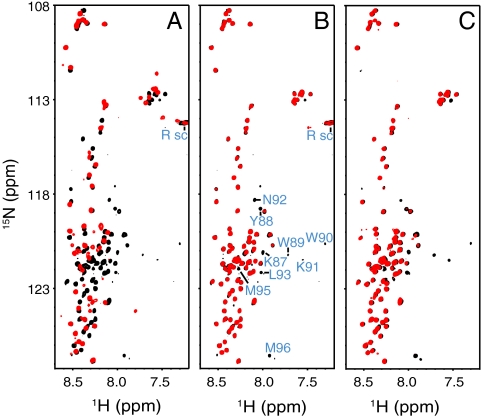
SNARE proteins play a critical role in intracellular membrane fusion by forming tight complexes that bring two membranes together and involve sequences called SNARE motifs. These motifs have a high tendency to form amphipathic coiled-coils that assemble into four-helix bundles, and often precede transmembrane regions. NMR studies in dodecylphosphocholine (DPC) micelles suggested that the N-terminal half of the SNARE motif from the neuronal SNARE synaptobrevin binds to membranes, which appeared to contradict previous biophysical studies of synaptobrevin in liposomes. NMR analyses of synaptobrevin reconstituted into nanodiscs and into liposomes now show that most of its SNARE motif, except for the basic C terminus, is highly flexible, exhibiting cross-peak patterns and transverse relaxation rates that are very similar to those observed in solution. Considering the proximity to the bilayer imposed by membrane anchoring, our data show that most of the synaptobrevin SNARE motif has a remarkable reluctance to bind membranes. This conclusion is further supported by NMR experiments showing that the soluble synaptobrevin SNARE motif does not bind to liposomes, even though it does bind to DPC micelles. These results show that nanodiscs provide a much better membrane model than DPC micelles in this system, and that most of the SNARE motif of membrane-anchored synaptobrevin is accessible for SNARE complex formation. We propose that the charge and hydrophobicity of SNARE motifs is optimized to enable formation of highly stable SNARE complexes while at the same time avoiding membrane binding, which could hinder SNARE complex assembly.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
