

Efficient isolation of Pseudomonas aeruginosa type III secretion translocators and assembly of heteromeric transmembrane pores in model membranes
- PMID: 21770428
- PMCID: PMC3171962
- DOI: 10.1021/bi200905x
Efficient isolation of Pseudomonas aeruginosa type III secretion translocators and assembly of heteromeric transmembrane pores in model membranes
Abstract
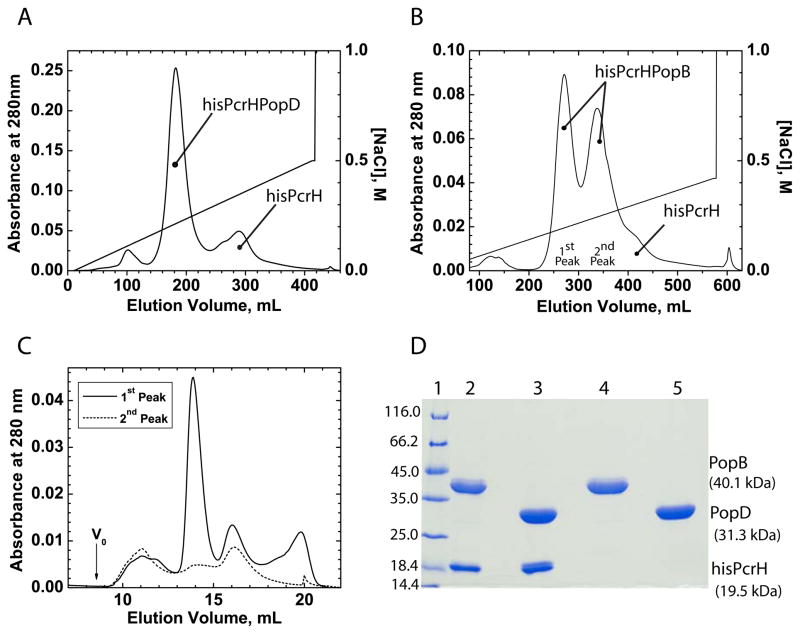
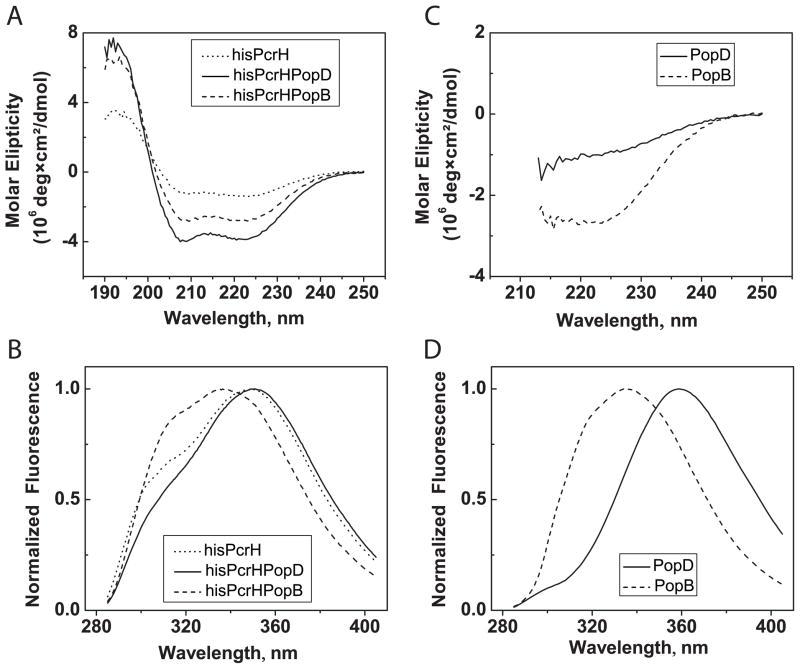
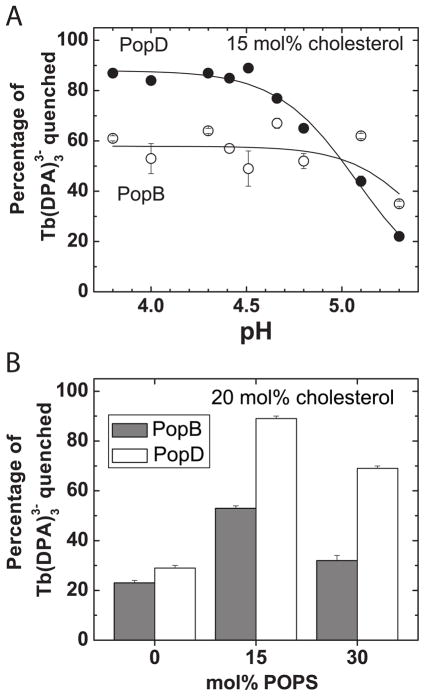
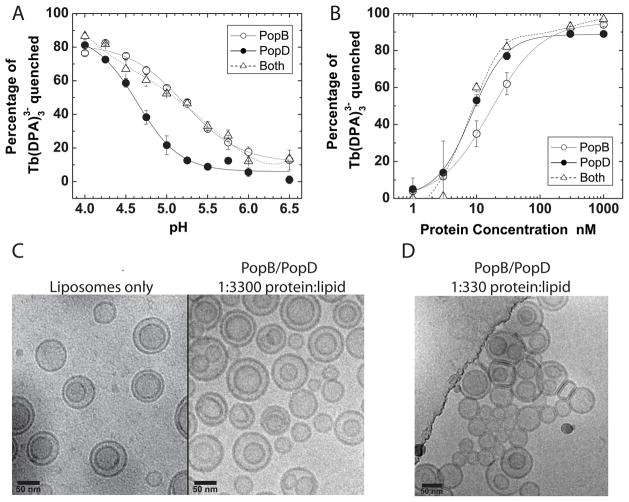
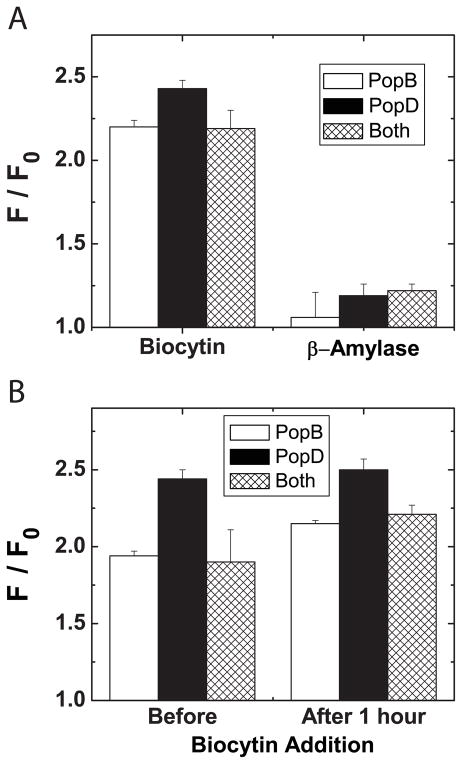
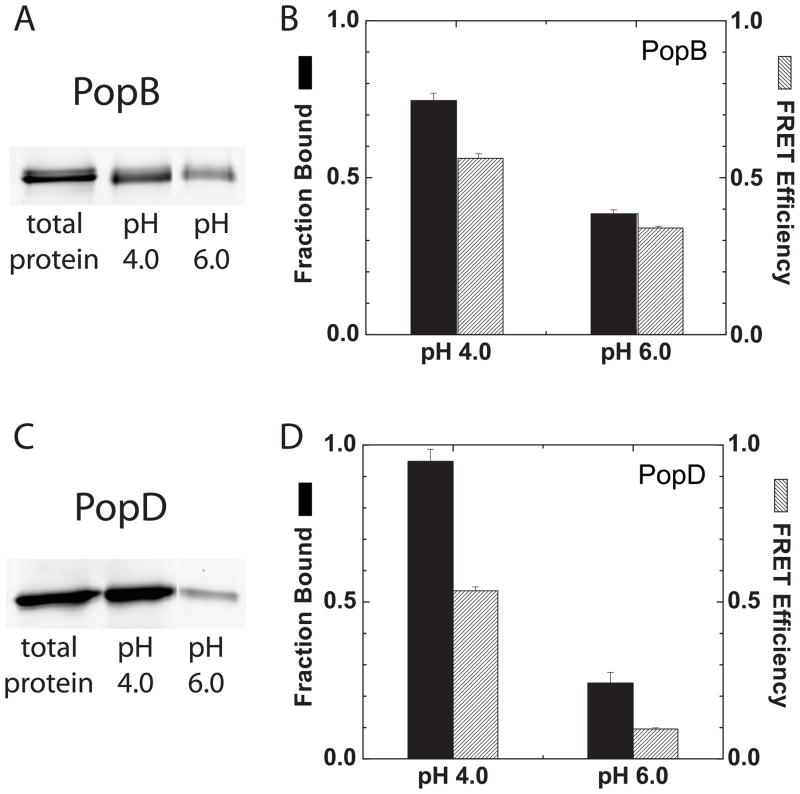
Translocation of bacterial toxins or effectors into host cells using the type III secretion (T3S) system is a conserved mechanism shared by many Gram-negative pathogens. Pseudomonas aeruginosa injects different proteins across the plasma membrane of target cells, altering the normal metabolism of the host. Protein translocation presumably occurs through a proteinaceous transmembrane pore formed by two T3S secreted protein translocators, PopB and PopD. Unfolded translocators are secreted through the T3S needle prior to insertion into the target membrane. Purified PopB and PopD form pores in model membranes. However, their tendency to form heterogeneous aggregates in solution had hampered the analysis of how these proteins undergo the transition from a denatured state to a membrane-inserted state. Translocators were purified as stable complexes with the cognate chaperone PcrH and isolated from the chaperone using 6 M urea. We report here the assembly of stable transmembrane pores by dilution of urea-denatured translocators in the presence of membranes. PopB and PopD spontaneously bound liposomes containing anionic phospholipids and cholesterol in a pH-dependent manner as observed by two independent assays, time-resolved Förster resonance energy transfer and sucrose-step gradient ultracentrifugation. Using Bodipy-labeled proteins, we found that PopB interacts with PopD on the membrane surface as determined by excitation energy migration and fluorescence quenching. Stable transmembrane pores are more efficiently assembled at pH <5.0, suggesting that acidic residues might be involved in the initial membrane binding and/or insertion. Altogether, the experimental setup described here represents an efficient method for the reconstitution and analysis of membrane-inserted translocators.
© 2011 American Chemical Society
Figures
References
-
- Galan JE, Wolf-Watz H. Protein delivery into eukaryotic cells by type III secretion machines. Nature. 2006;444:567–573. - PubMed
-
- Cornelis GR. The type III secretion injectisome. Nat Rev Micro. 2006;4:811–825. - PubMed
-
- Frank D. The exoenzyme S regulon of Pseudomonas aeruginosa. Mol Microbiol. 1997;26:621–629. - PubMed
-
- Hauser AR, Cobb E, Bodi M, Mariscal D, Valles J, Engel JN, Rello J. Type III protein secretion is associated with poor clinical outcomes in patients with ventilator-associated pneumonia caused by Pseudomonas aeruginosa. Crit Care Med. 2002;30:521–528. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
