

Arrested oocyst maturation in Plasmodium parasites lacking type II NADH:ubiquinone dehydrogenase
- PMID: 21771793
- PMCID: PMC3173203
- DOI: 10.1074/jbc.M111.269399
Arrested oocyst maturation in Plasmodium parasites lacking type II NADH:ubiquinone dehydrogenase
Abstract
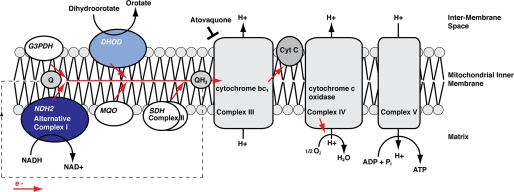
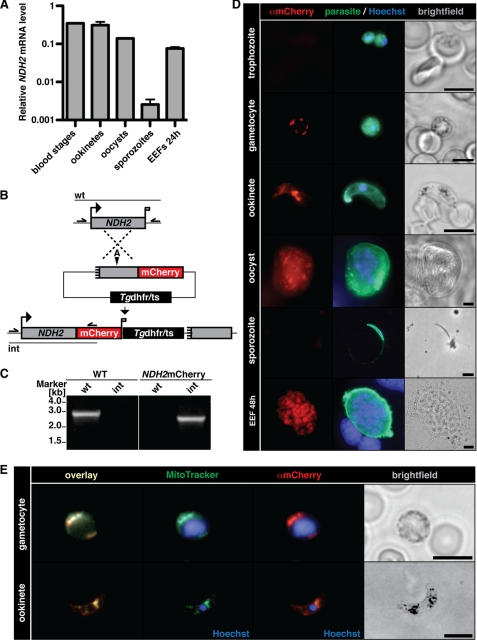
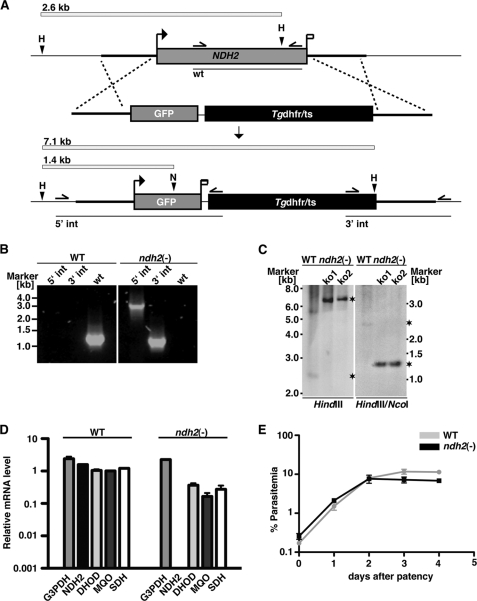
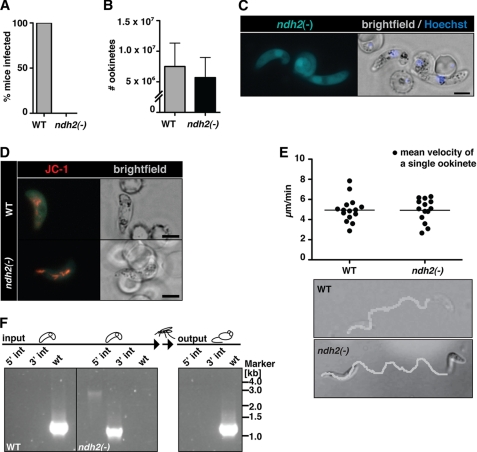
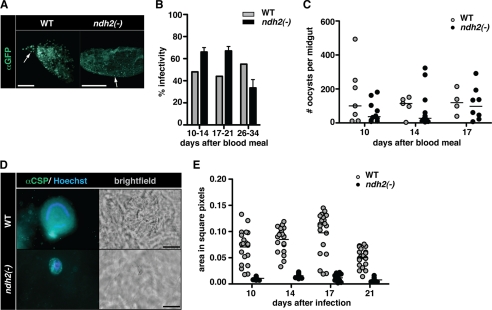
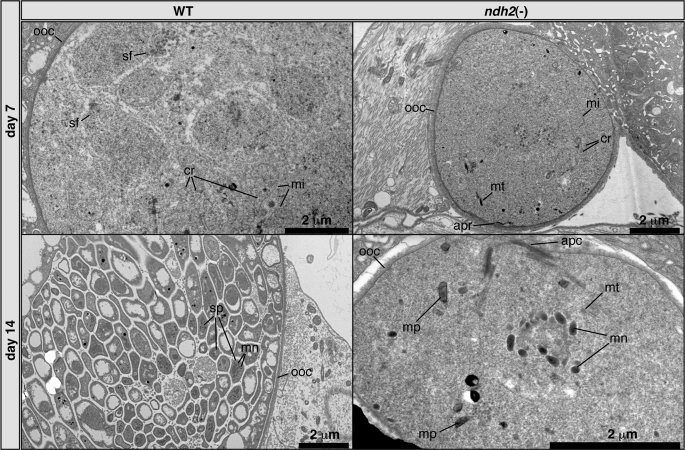
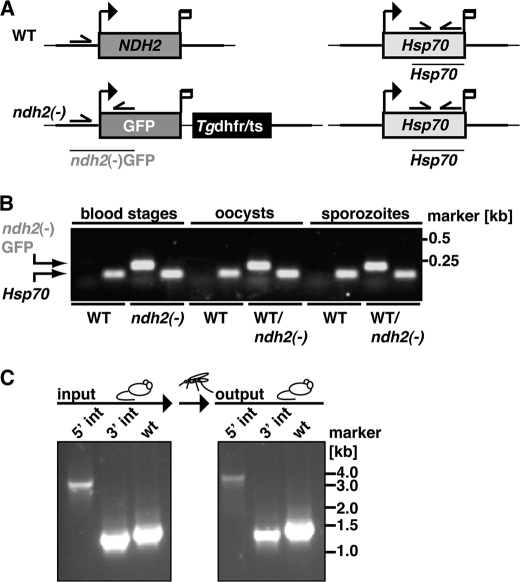
The Plasmodium mitochondrial electron transport chain has received considerable attention as a potential target for new antimalarial drugs. Atovaquone, a potent inhibitor of Plasmodium cytochrome bc(1), in combination with proguanil is recommended for chemoprophylaxis and treatment of malaria. The type II NADH:ubiquinone oxidoreductase (NDH2) is considered an attractive drug target, as its inhibition is thought to lead to the arrest of the mitochondrial electron transport chain and, as a consequence, pyrimidine biosynthesis, an essential pathway for the parasite. Using the rodent malaria parasite Plasmodium berghei as an in vivo infection model, we studied the role of NDH2 during Plasmodium life cycle progression. NDH2 can be deleted by targeted gene disruption and, thus, is dispensable for the pathogenic asexual blood stages, disproving the candidacy for an anti-malarial drug target. After transmission to the insect vector, NDH2-deficient ookinetes display an intact mitochondrial membrane potential. However, ndh2(-) parasites fail to develop into mature oocysts in the mosquito midgut. We propose that Plasmodium blood stage parasites rely on glycolysis as the main ATP generating process, whereas in the invertebrate vector, a glucose-deprived environment, the malaria parasite is dependent on an intact mitochondrial respiratory chain.
Figures
References
-
- World Health Organization (2010) World Malaria Report 2010, World Health Organization Press, Geneva, Switzerland
-
- World Health Organization (2010) Global Report on Antimalarial Drug Efficacy and Drug Resistance: 2000–2010, World Health Organization Press, Geneva, Switzerland
-
- Petersen I., Eastman R., Lanzer M. (2011) FEBS Lett. 585, 1551–1562 - PubMed
-
- Painter H. J., Morrisey J. M., Mather M. W., Vaidya A. B. (2007) Nature 446, 88–91 - PubMed
-
- Mather M. W., Vaidya A. B. (2008) J. Bioenerg. Biomembr. 40, 425–433 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
