

Notch and Ras promote sequential steps of excretory tube development in C. elegans
- PMID: 21771815
- PMCID: PMC3143567
- DOI: 10.1242/dev.068148
Notch and Ras promote sequential steps of excretory tube development in C. elegans
Abstract
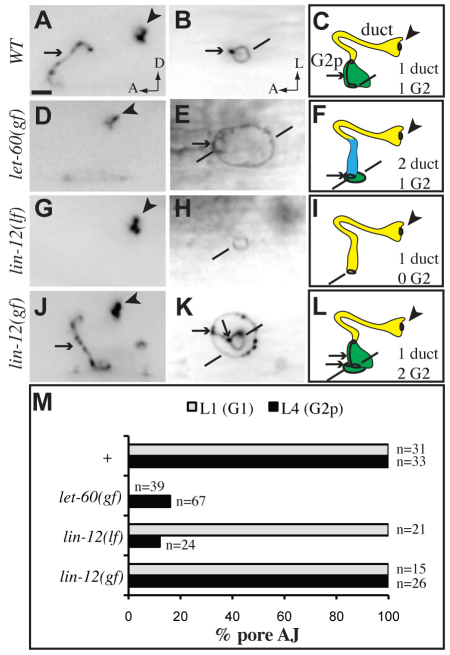
Receptor tyrosine kinases and Notch are crucial for tube formation and branching morphogenesis in many systems, but the specific cellular processes that require signaling are poorly understood. Here we describe sequential roles for Notch and Epidermal growth factor (EGF)-Ras-ERK signaling in the development of epithelial tube cells in the C. elegans excretory (renal-like) organ. This simple organ consists of three tandemly connected unicellular tubes: the excretory canal cell, duct and G1 pore. lin-12 and glp-1/Notch are required to generate the canal cell, which is a source of LIN-3/EGF ligand and physically attaches to the duct during de novo epithelialization and tubulogenesis. Canal cell asymmetry and let-60/Ras signaling influence which of two equivalent precursors will attach to the canal cell. Ras then specifies duct identity, inducing auto-fusion and a permanent epithelial character; the remaining precursor becomes the G1 pore, which eventually loses epithelial character and withdraws from the organ to become a neuroblast. Ras continues to promote subsequent aspects of duct morphogenesis and differentiation, and acts primarily through Raf-ERK and the transcriptional effectors LIN-1/Ets and EOR-1. These results reveal multiple genetically separable roles for Ras signaling in tube development, as well as similarities to Ras-mediated control of branching morphogenesis in more complex organs, including the mammalian kidney. The relative simplicity of the excretory system makes it an attractive model for addressing basic questions about how cells gain or lose epithelial character and organize into tubular networks.
Figures






Similar articles
-
The Lipocalin LPR-1 Cooperates with LIN-3/EGF Signaling To Maintain Narrow Tube Integrity in Caenorhabditis elegans.Genetics. 2017 Mar;205(3):1247-1260. doi: 10.1534/genetics.116.195156. Epub 2016 Dec 30. Genetics. 2017. PMID: 28040739 Free PMC article.
-
A non-cell-autonomous role for Ras signaling in C. elegans neuroblast delamination.Development. 2014 Nov;141(22):4279-84. doi: 10.1242/dev.112045. Development. 2014. PMID: 25371363 Free PMC article.
-
RTK/Ras/MAPK signaling.WormBook. 2006 Feb 11:1-19. doi: 10.1895/wormbook.1.80.1. WormBook. 2006. PMID: 18050474 Free PMC article. Review.
-
lin-1 has both positive and negative functions in specifying multiple cell fates induced by Ras/MAP kinase signaling in C. elegans.Dev Biol. 2005 Oct 1;286(1):338-51. doi: 10.1016/j.ydbio.2005.08.007. Dev Biol. 2005. PMID: 16140291
-
Vulval development.WormBook. 2005 Jun 25:1-28. doi: 10.1895/wormbook.1.6.1. WormBook. 2005. PMID: 18050418 Free PMC article. Review.
Cited by
-
A survey of the kinome pharmacopeia reveals multiple scaffolds and targets for the development of novel anthelmintics.Sci Rep. 2021 Apr 28;11(1):9161. doi: 10.1038/s41598-021-88150-6. Sci Rep. 2021. PMID: 33911106 Free PMC article.
-
Gap junctions in C. elegans: Their roles in behavior and development.Dev Neurobiol. 2017 May;77(5):587-596. doi: 10.1002/dneu.22408. Epub 2016 Jun 22. Dev Neurobiol. 2017. PMID: 27294317 Free PMC article. Review.
-
The Double-Edged Proteins in Cancer Proteomes and the Generation of Induced Tumor-Suppressing Cells (iTSCs).Proteomes. 2023 Jan 18;11(1):5. doi: 10.3390/proteomes11010005. Proteomes. 2023. PMID: 36810561 Free PMC article. Review.
-
A Receptor Tyrosine Kinase Network Regulates Neuromuscular Function in Response to Oxidative Stress in Caenorhabditis elegans.Genetics. 2019 Apr;211(4):1283-1295. doi: 10.1534/genetics.119.302026. Epub 2019 Feb 19. Genetics. 2019. PMID: 30782598 Free PMC article.
-
Quantitative Differences in Nuclear β-catenin and TCF Pattern Embryonic Cells in C. elegans.PLoS Genet. 2015 Oct 21;11(10):e1005585. doi: 10.1371/journal.pgen.1005585. eCollection 2015 Oct. PLoS Genet. 2015. PMID: 26488501 Free PMC article.
References
-
- Bar T., Guldner F. H., Wolff J. R. (1984). `Seamless' endothelial cells of blood capillaries. Cell Tissue Res. 235, 99-106 - PubMed
-
- Beitel G. J., Tuck S., Greenwald I., Horvitz H. R. (1995). The Caenorhabditis elegans gene lin-1 encodes an ETS-domain protein and defines a branch of the vulval induction pathway. Genes Dev. 9, 3149-3162 - PubMed
-
- Berry K. L., Bulow H. E., Hall D. H., Hobert O. (2003). A C. elegans CLIC-like protein required for intracellular tube formation and maintenance. Science 302, 2134-2137 - PubMed
-
- Bossinger O., Klebes A., Segbert C., Theres C., Knust E. (2001). Zonula adherens formation in Caenorhabditis elegans requires dlg-1, the homologue of the Drosophila gene discs large. Dev. Biol. 230, 29-42 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
