

A Sox10 enhancer element common to the otic placode and neural crest is activated by tissue-specific paralogs
- PMID: 21775416
- PMCID: PMC3152925
- DOI: 10.1242/dev.057836
A Sox10 enhancer element common to the otic placode and neural crest is activated by tissue-specific paralogs
Abstract
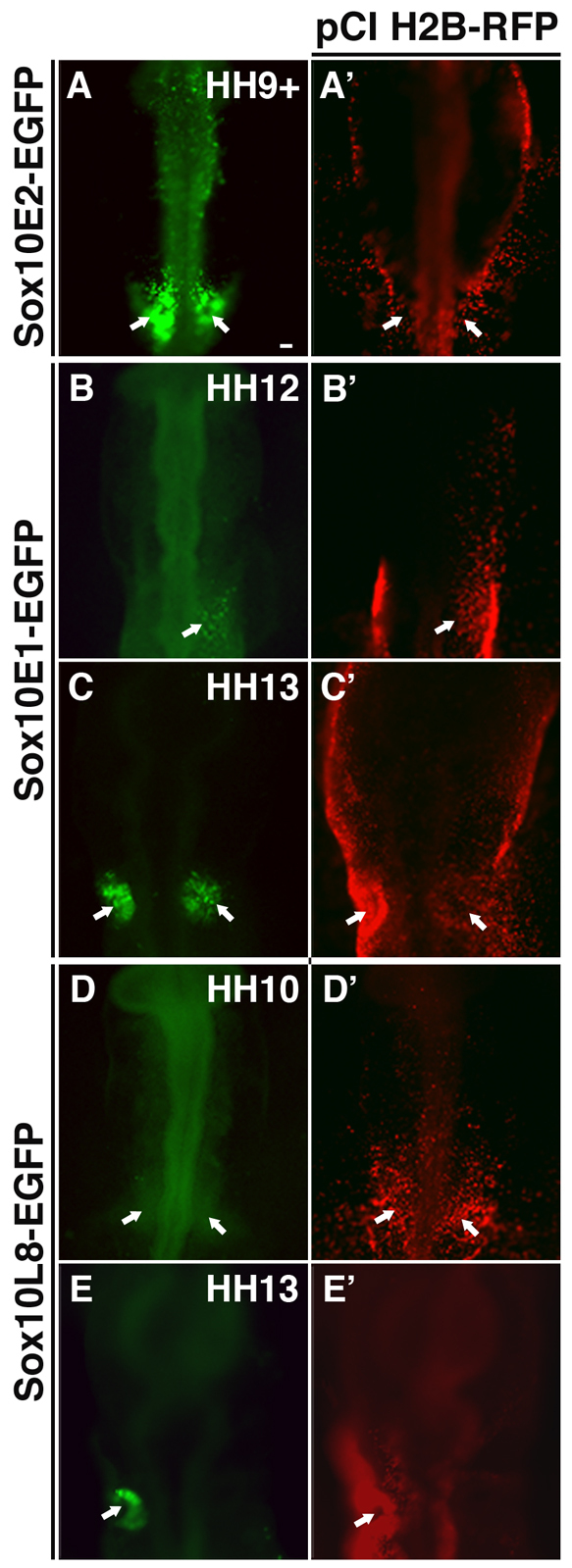
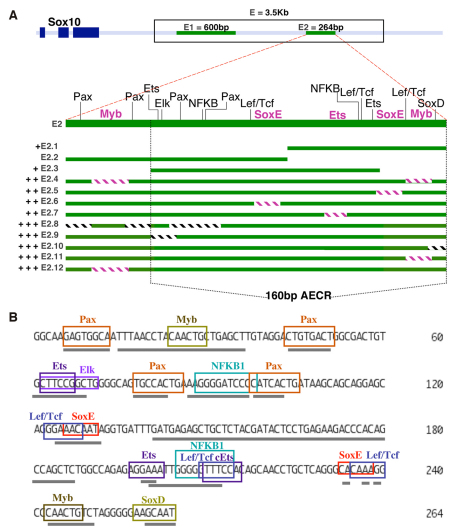
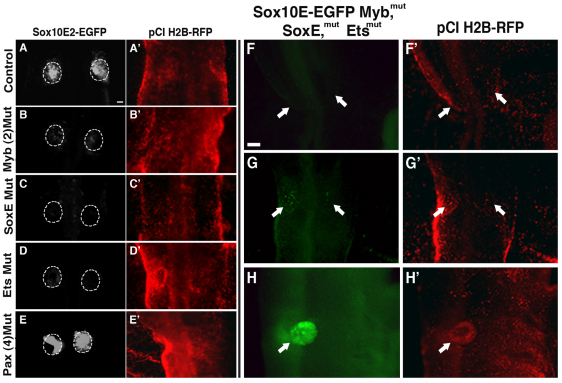
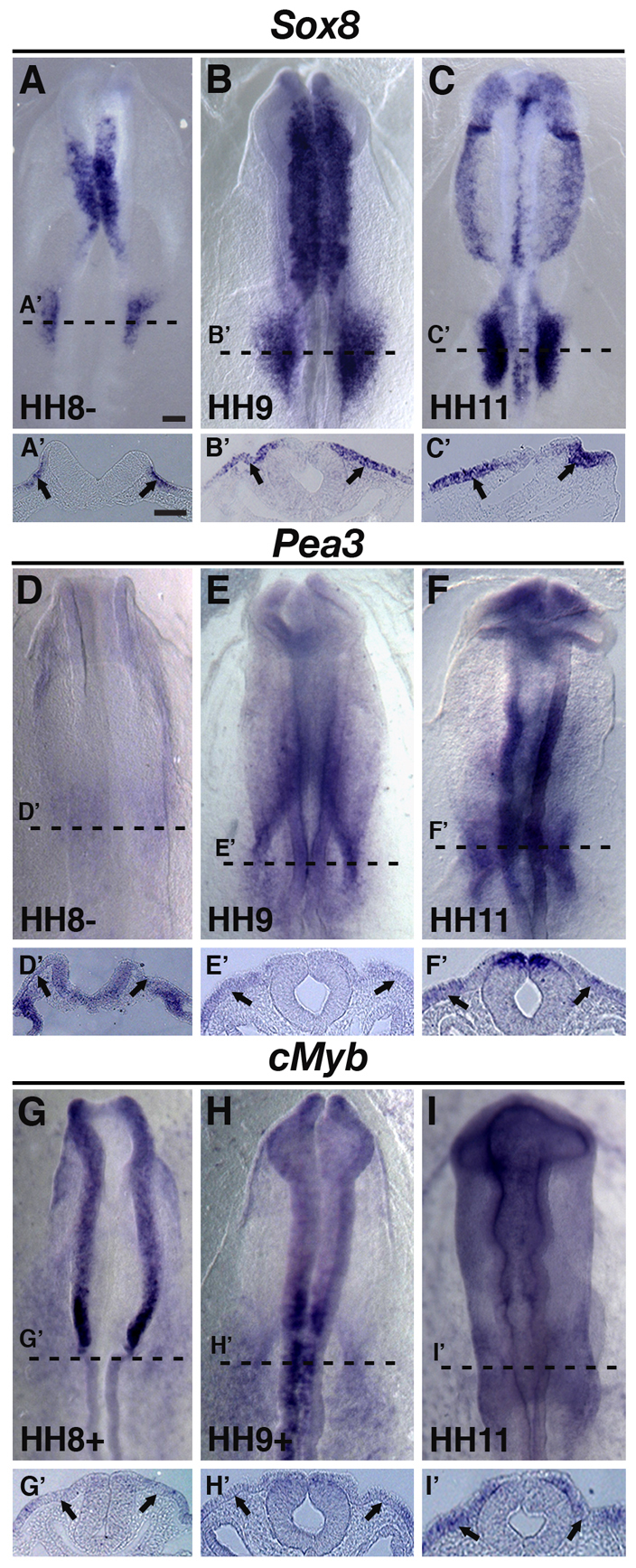
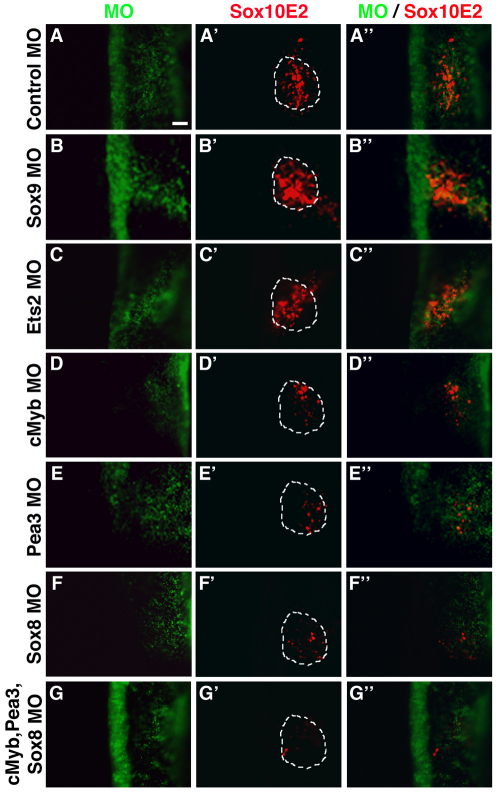
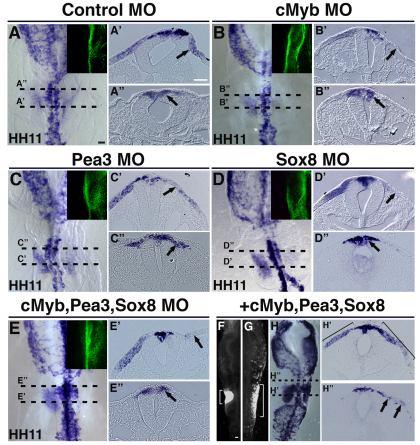
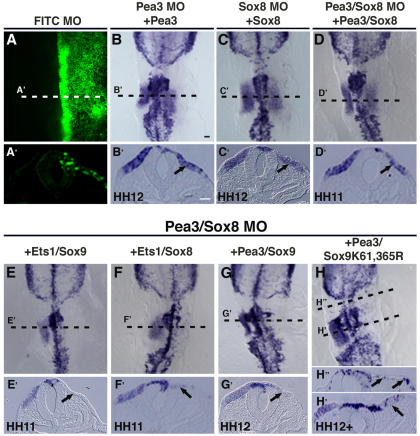
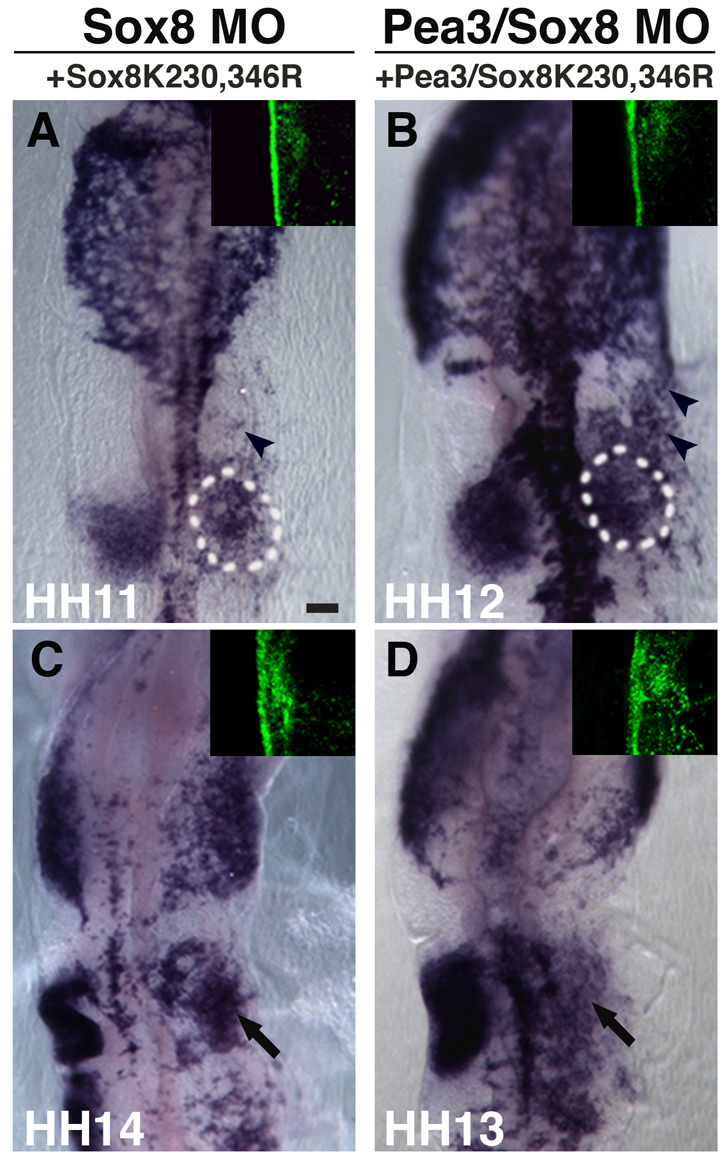
The otic placode, a specialized region of ectoderm, gives rise to components of the inner ear and shares many characteristics with the neural crest, including expression of the key transcription factor Sox10. Here, we show that in avian embryos, a highly conserved cranial neural crest enhancer, Sox10E2, also controls the onset of Sox10 expression in the otic placode. Interestingly, we show that different combinations of paralogous transcription factors (Sox8, Pea3 and cMyb versus Sox9, Ets1 and cMyb) are required to mediate Sox10E2 activity in the ear and neural crest, respectively. Mutating their binding motifs within Sox10E2 greatly reduces enhancer activity in the ear. Moreover, simultaneous knockdown of Sox8, Pea3 and cMyb eliminates not only the enhancer-driven reporter expression, but also the onset of endogenous Sox10 expression in the ear. Rescue experiments confirm that the specific combination of Myb together with Sox8 and Pea3 is responsible for the onset of Sox10 expression in the otic placode, as opposed to Myb plus Sox9 and Ets1 for neural crest Sox10 expression. Whereas SUMOylation of Sox8 is not required for the initial onset of Sox10 expression, it is necessary for later otic vesicle formation. This new role of Sox8, Pea3 and cMyb in controlling Sox10 expression via a common otic/neural crest enhancer suggests an evolutionarily conserved function for the combination of paralogous transcription factors in these tissues of distinct embryological origin.
Figures
References
-
- Bagheri-Fam S., Barrionuevo F., Dohrmann U., Gunther T., Schule R., Kemler R., Mallo M., Kanzler B., Scherer G. (2006). Long-range upstream and downstream enhancers control distinct subsets of the complex spatiotemporal Sox9 expression pattern. Dev. Biol. 291, 382-397 - PubMed
-
- Baker C. V. (2008). The evolution and elaboration of vertebrate neural crest cells. Curr. Opin. Genet. Dev. 18, 536-543 - PubMed
-
- Barrionuevo F., Naumann A., Bagheri-Fam S., Speth V., Taketo M. M., Scherer G., Neubuser A. (2008). Sox9 is required for invagination of the otic placode in mice. Dev. Biol. 317, 213-224 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
