

A conserved long noncoding RNA affects sleep behavior in Drosophila
- PMID: 21775470
- PMCID: PMC3189806
- DOI: 10.1534/genetics.111.131706
A conserved long noncoding RNA affects sleep behavior in Drosophila
Abstract
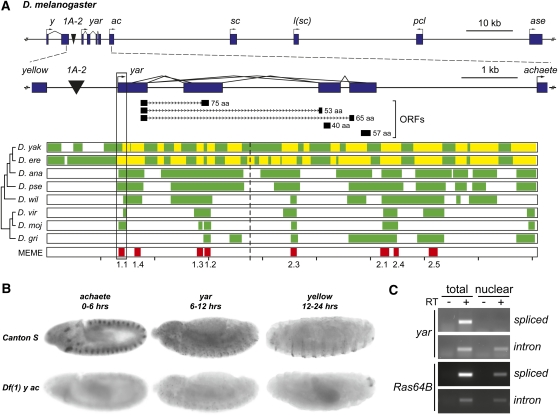
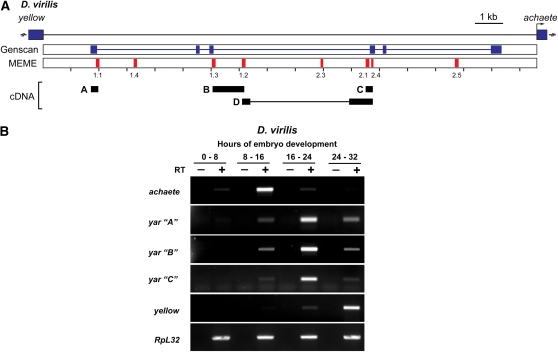
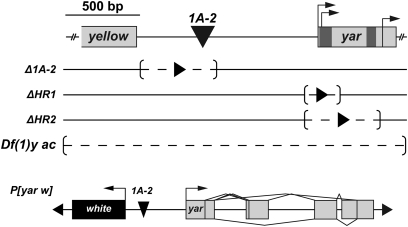
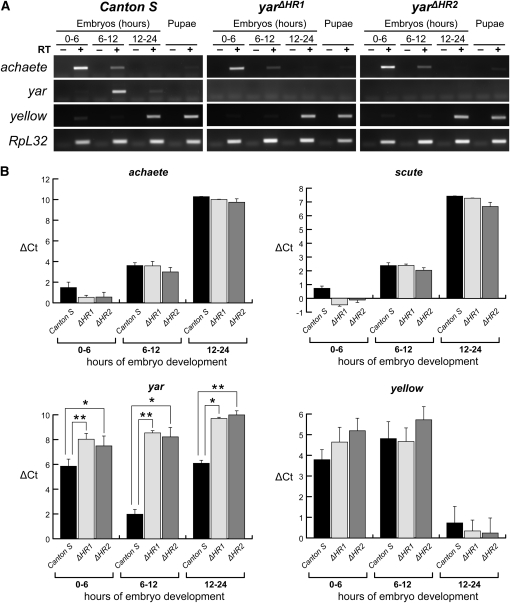
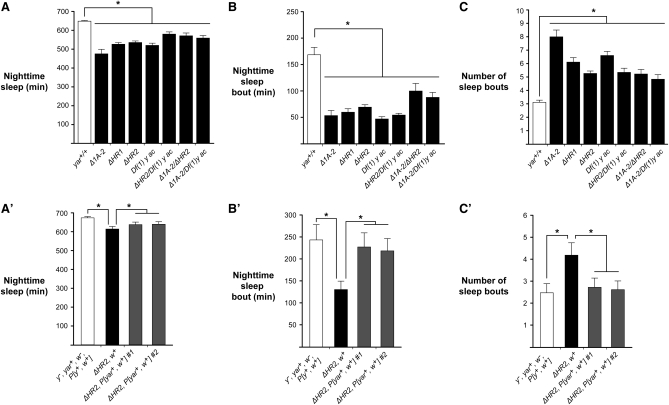
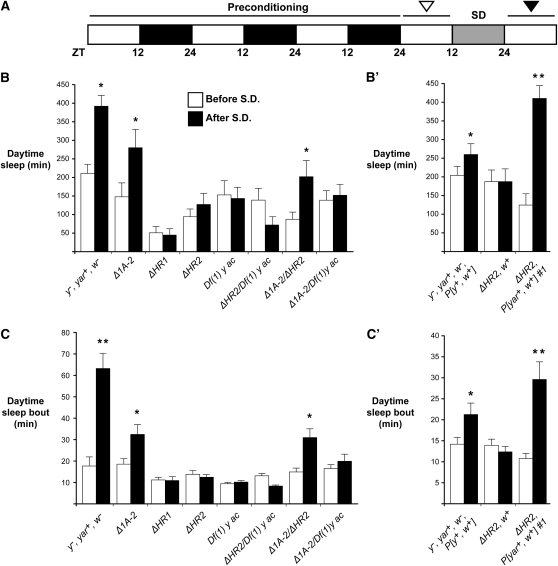
Metazoan genomes encode an abundant collection of mRNA-like, long noncoding (lnc)RNAs. Although lncRNAs greatly expand the transcriptional repertoire, we have a limited understanding of how these RNAs contribute to developmental regulation. Here, we investigate the function of the Drosophila lncRNA called yellow-achaete intergenic RNA (yar). Comparative sequence analyses show that the yar gene is conserved in Drosophila species representing 40-60 million years of evolution, with one of the conserved sequence motifs encompassing the yar promoter. Further, the timing of yar expression in Drosophila virilis parallels that in D. melanogaster, suggesting that transcriptional regulation of yar is conserved. The function of yar was defined by generating null alleles. Flies lacking yar RNAs are viable and show no overt morphological defects, consistent with maintained transcriptional regulation of the adjacent yellow (y) and achaete (ac) genes. The location of yar within a neural gene cluster led to the investigation of effects of yar in behavioral assays. These studies demonstrated that loss of yar alters sleep regulation in the context of a normal circadian rhythm. Nighttime sleep was reduced and fragmented, with yar mutants displaying diminished sleep rebound following sleep deprivation. Importantly, these defects were rescued by a yar transgene. These data provide the first example of a lncRNA gene involved in Drosophila sleep regulation. We find that yar is a cytoplasmic lncRNA, suggesting that yar may regulate sleep by affecting stabilization or translational regulation of mRNAs. Such functions of lncRNAs may extend to vertebrates, as lncRNAs are abundant in neural tissues.
Figures
References
-
- Anaka M., MacDonald C. D., Barkova E., Simon K., Rostom R., et al. , 2008. The white gene of Drosophila melanogaster encodes a protein with a role in courtship behavior. J. Neurogenet. 22: 243–276 - PubMed
-
- Barrandon C., Spiluttini B., Bensaude O., 2008. Non-coding RNAs regulating the transcriptional machinery. Biol. Cell 100: 83–95 - PubMed
-
- Brock H. W., Hodgson J. W., Petruk S., Mazo A., 2009. Regulatory noncoding RNAs at Hox loci. Biochem. Cell Biol. 87: 27–34 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
