

RNA-ligase-dependent biases in miRNA representation in deep-sequenced small RNA cDNA libraries
- PMID: 21775473
- PMCID: PMC3162335
- DOI: 10.1261/rna.2799511
RNA-ligase-dependent biases in miRNA representation in deep-sequenced small RNA cDNA libraries
Abstract
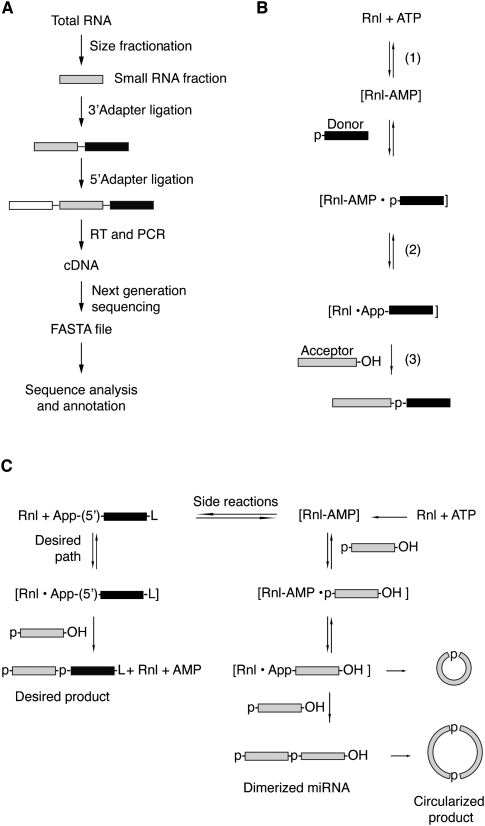
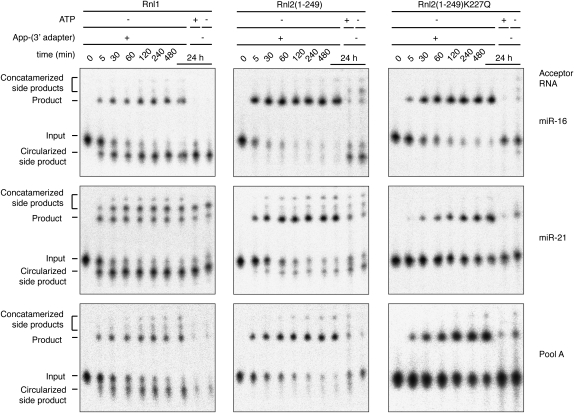
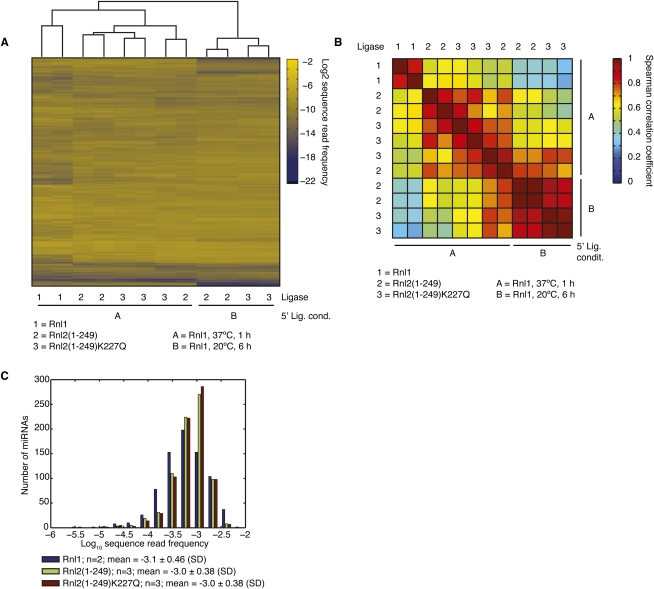
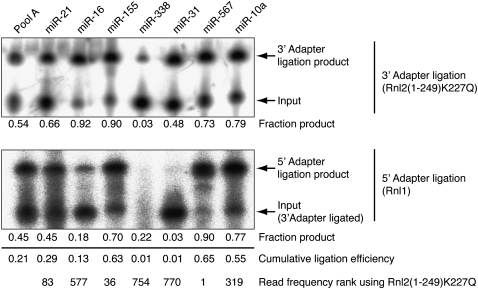
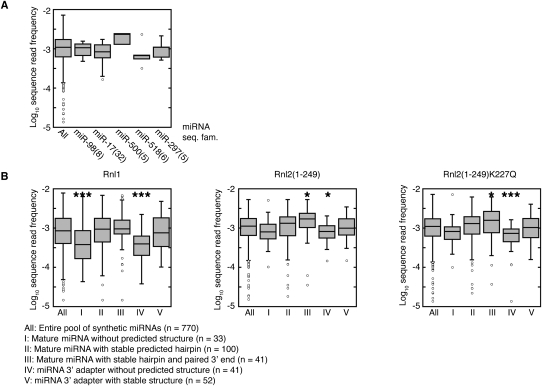
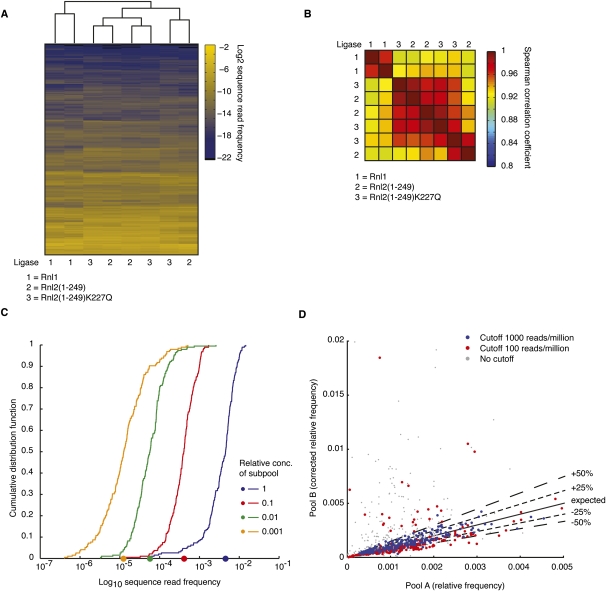
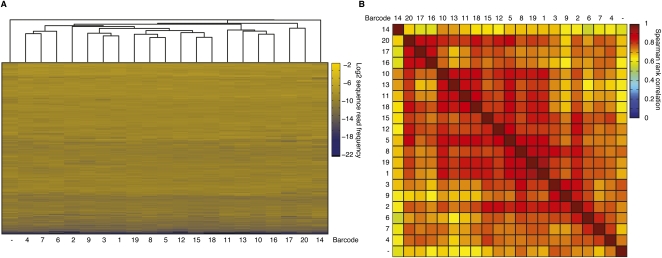
Sequencing of small RNA cDNA libraries is an important tool for the discovery of new RNAs and the analysis of their mutational status as well as expression changes across samples. It requires multiple enzyme-catalyzed steps, including sequential oligonucleotide adapter ligations to the 3' and 5' ends of the small RNAs, reverse transcription (RT), and PCR. We assessed biases in representation of miRNAs relative to their input concentration, using a pool of 770 synthetic miRNAs and 45 calibrator oligoribonucleotides, and tested the influence of Rnl1 and two variants of Rnl2, Rnl2(1-249) and Rnl2(1-249)K227Q, for 3'-adapter ligation. The use of the Rnl2 variants for adapter ligations yielded substantially fewer side products compared with Rnl1; however, the benefits of using Rnl2 remained largely obscured by additional biases in the 5'-adapter ligation step; RT and PCR steps did not have a significant impact on read frequencies. Intramolecular secondary structures of miRNA and/or miRNA/3'-adapter products contributed to these biases, which were highly reproducible under defined experimental conditions. We used the synthetic miRNA cocktail to derive correction factors for approximation of the absolute levels of individual miRNAs in biological samples. Finally, we evaluated the influence of 5'-terminal 5-nt barcode extensions for a set of 20 barcoded 3' adapters and observed similar biases in miRNA read distribution, thereby enabling cost-saving multiplex analysis for large-scale miRNA profiling.
Figures
Similar articles
-
Structural bias in T4 RNA ligase-mediated 3'-adapter ligation.Nucleic Acids Res. 2012 Apr;40(7):e54. doi: 10.1093/nar/gkr1263. Epub 2012 Jan 12. Nucleic Acids Res. 2012. PMID: 22241775 Free PMC article.
-
Highly efficient ligation of small RNA molecules for microRNA quantitation by high-throughput sequencing.J Vis Exp. 2014 Nov 18;(93):e52095. doi: 10.3791/52095. J Vis Exp. 2014. PMID: 25490151 Free PMC article.
-
T4 RNA ligase 2 truncated active site mutants: improved tools for RNA analysis.BMC Biotechnol. 2011 Jul 1;11:72. doi: 10.1186/1472-6750-11-72. BMC Biotechnol. 2011. PMID: 21722378 Free PMC article.
-
MicroRNA (miRNA) Profiling.Methods Mol Biol. 2016;1381:151-61. doi: 10.1007/978-1-4939-3204-7_8. Methods Mol Biol. 2016. PMID: 26667459 Review.
-
Biases in small RNA deep sequencing data.Nucleic Acids Res. 2014 Feb;42(3):1414-26. doi: 10.1093/nar/gkt1021. Epub 2013 Nov 5. Nucleic Acids Res. 2014. PMID: 24198247 Free PMC article. Review.
Cited by
-
Deep sequencing reveals predominant expression of miR-21 amongst the small non-coding RNAs in retinal microvascular endothelial cells.J Cell Biochem. 2012 Jun;113(6):2098-111. doi: 10.1002/jcb.24084. J Cell Biochem. 2012. PMID: 22298343 Free PMC article.
-
MicroRNAs in human cancer.Adv Exp Med Biol. 2013;774:1-20. doi: 10.1007/978-94-007-5590-1_1. Adv Exp Med Biol. 2013. PMID: 23377965 Free PMC article. Review.
-
exRNA Atlas Analysis Reveals Distinct Extracellular RNA Cargo Types and Their Carriers Present across Human Biofluids.Cell. 2019 Apr 4;177(2):463-477.e15. doi: 10.1016/j.cell.2019.02.018. Cell. 2019. PMID: 30951672 Free PMC article.
-
Evaluating and Correcting Inherent Bias of microRNA Expression in Illumina Sequencing Analysis.Front Mol Biosci. 2019 Apr 24;6:17. doi: 10.3389/fmolb.2019.00017. eCollection 2019. Front Mol Biosci. 2019. PMID: 31069233 Free PMC article.
-
High-efficiency RNA cloning enables accurate quantification of miRNA expression by deep sequencing.Genome Biol. 2013;14(10):R109. doi: 10.1186/gb-2013-14-10-r109. Genome Biol. 2013. PMID: 24098942 Free PMC article.
References
-
- Aravin AA, Tuschl T 2005. Identification and characterization of small RNAs involved in RNA silencing. FEBS Lett 579: 5830–5840 - PubMed
-
- Berninger P, Gaidatzis D, van Nimwegen E, Zavolan M 2008. Computational analysis of small RNA cloning data. Methods 44: 13–21 - PubMed
-
- Bhattacharyya SN, Filipowicz W 2007. Argonautes and company: Sailing against the wind. Cell 128: 1027–1028 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials