

Postnatal loss of Dlk1 imprinting in stem cells and niche astrocytes regulates neurogenesis
- PMID: 21776083
- PMCID: PMC3160481
- DOI: 10.1038/nature10229
Postnatal loss of Dlk1 imprinting in stem cells and niche astrocytes regulates neurogenesis
Abstract
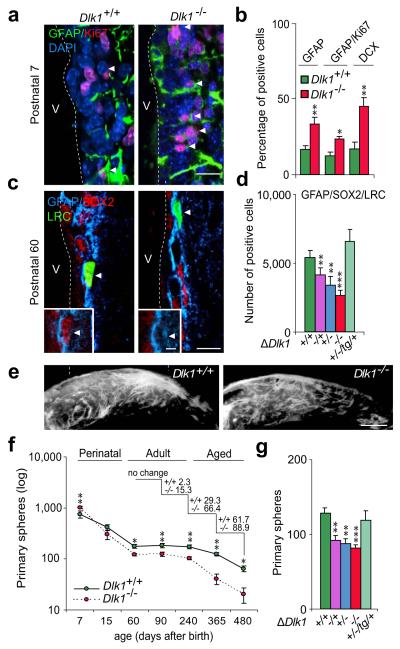
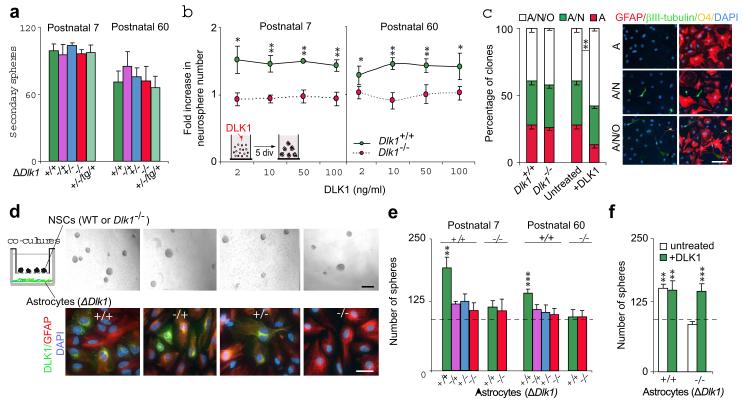
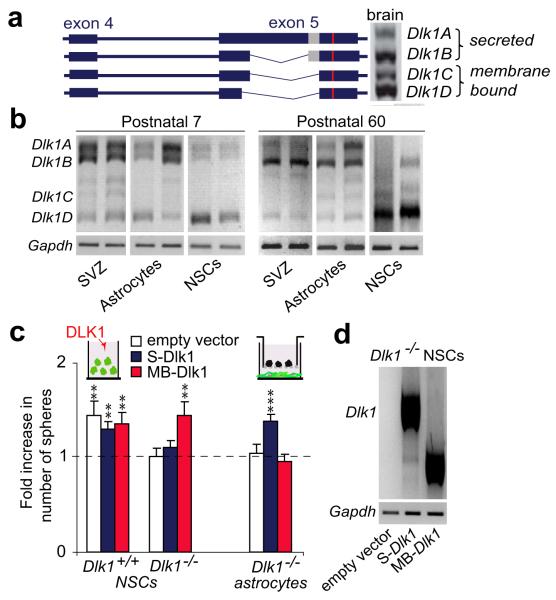
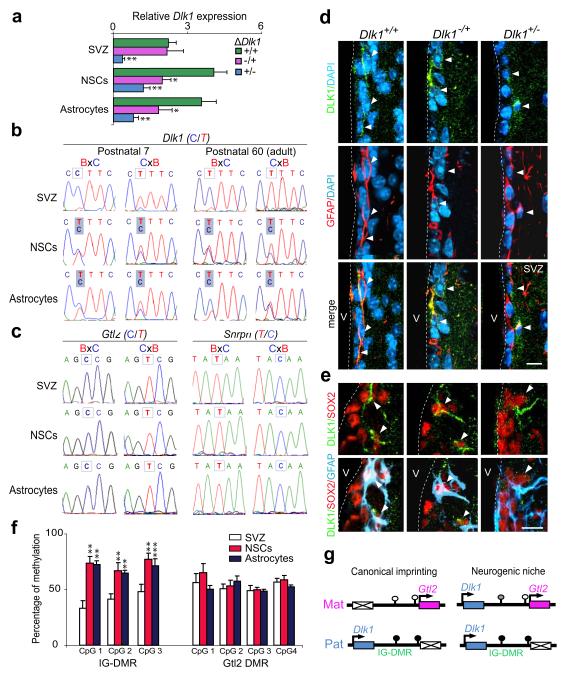
The gene for the atypical NOTCH ligand delta-like homologue 1 (Dlk1) encodes membrane-bound and secreted isoforms that function in several developmental processes in vitro and in vivo. Dlk1, a member of a cluster of imprinted genes, is expressed from the paternally inherited chromosome. Here we show that mice that are deficient in Dlk1 have defects in postnatal neurogenesis in the subventricular zone: a developmental continuum that results in depletion of mature neurons in the olfactory bulb. We show that DLK1 is secreted by niche astrocytes, whereas its membrane-bound isoform is present in neural stem cells (NSCs) and is required for the inductive effect of secreted DLK1 on self-renewal. Notably, we find that there is a requirement for Dlk1 to be expressed from both maternally and paternally inherited chromosomes. Selective absence of Dlk1 imprinting in both NSCs and niche astrocytes is associated with postnatal acquisition of DNA methylation at the germ-line-derived imprinting control region. The results emphasize molecular relationships between NSCs and the niche astrocyte cells of the microenvironment, identifying a signalling system encoded by a single gene that functions coordinately in both cell types. The modulation of genomic imprinting in a stem-cell environment adds a new level of epigenetic regulation to the establishment and maintenance of the niche, raising wider questions about the adaptability, function and evolution of imprinting in specific developmental contexts.
Figures
Comment in
-
Neuroscience: Imprinting in the brain.Nature. 2011 Jul 20;475(7356):299-300. doi: 10.1038/475299a. Nature. 2011. PMID: 21776070 Free PMC article.
-
Stem cells: a niche role for DLK1.Nat Rev Neurosci. 2011 Aug 10;12(9):489. doi: 10.1038/nrn3092. Nat Rev Neurosci. 2011. PMID: 21829221 No abstract available.
References
METHODS REFERENCES
-
- Lie DC, et al. Wnt signalling regulates adult hippocampal neurogenesis. Nature. 2005;437(7063):1370–1375. - PubMed
References
-
- Takada S, et al. Delta-like and Gtl2 are reciprocally expressed, differentially methylated linked imprinted genes on mouse chromosome 12. Curr Biol. 2000;10(18):1135–1138. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
