

β-myosin heavy chain is induced by pressure overload in a minor subpopulation of smaller mouse cardiac myocytes
- PMID: 21778428
- PMCID: PMC3166391
- DOI: 10.1161/CIRCRESAHA.111.243410
β-myosin heavy chain is induced by pressure overload in a minor subpopulation of smaller mouse cardiac myocytes
Abstract
Rationale: Induction of the fetal hypertrophic marker gene β-myosin heavy chain (β-MyHC) is a signature feature of pressure overload hypertrophy in rodents. β-MyHC is assumed present in all or most enlarged myocytes.
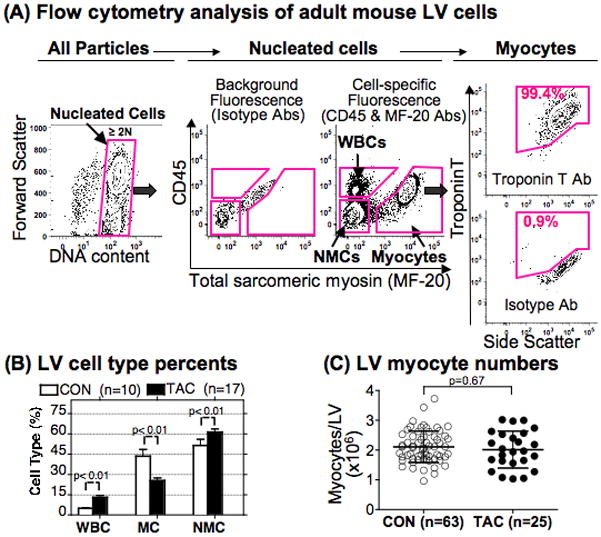
Objective: To quantify the number and size of myocytes expressing endogenous β-MyHC by a flow cytometry approach.
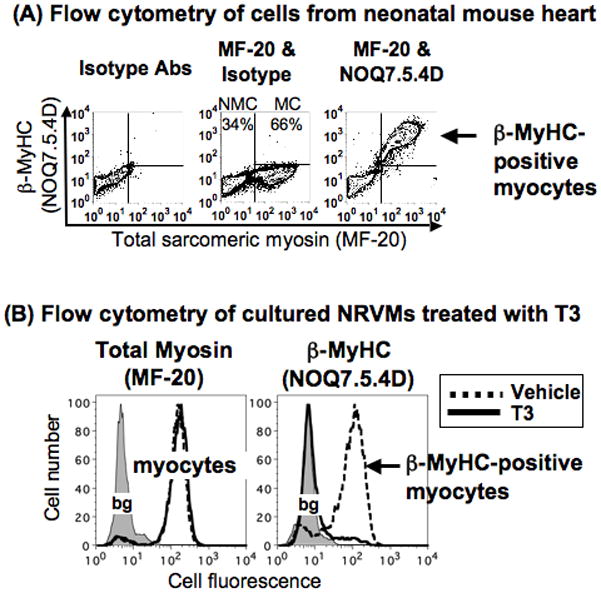
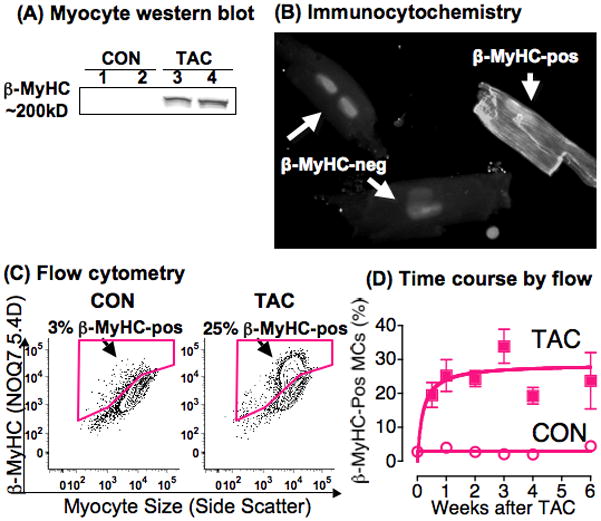
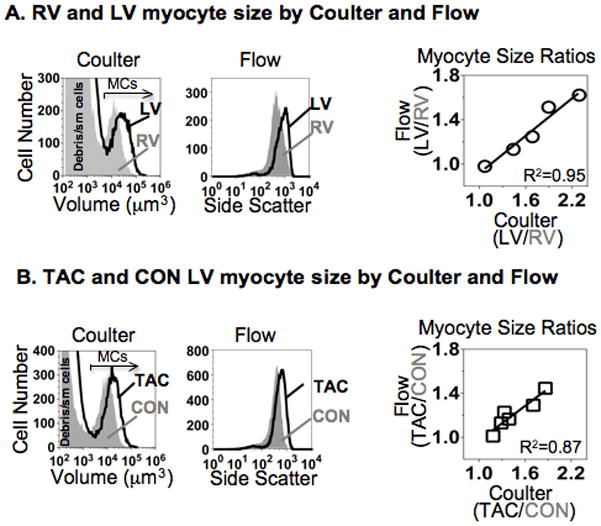
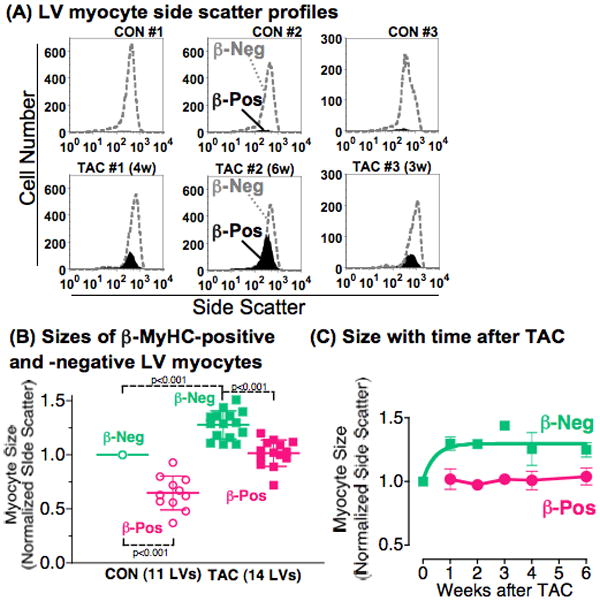
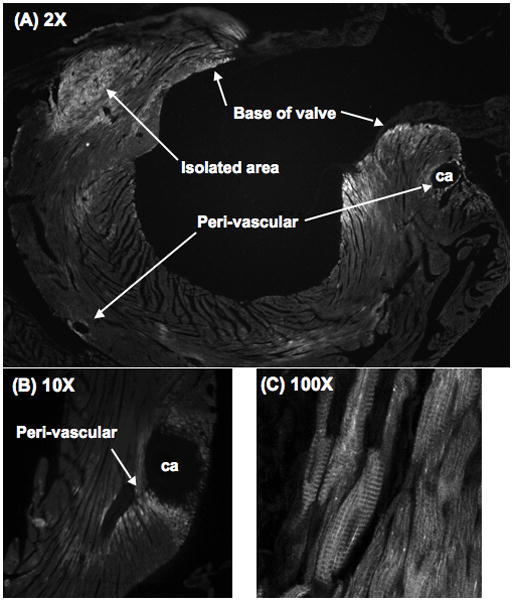
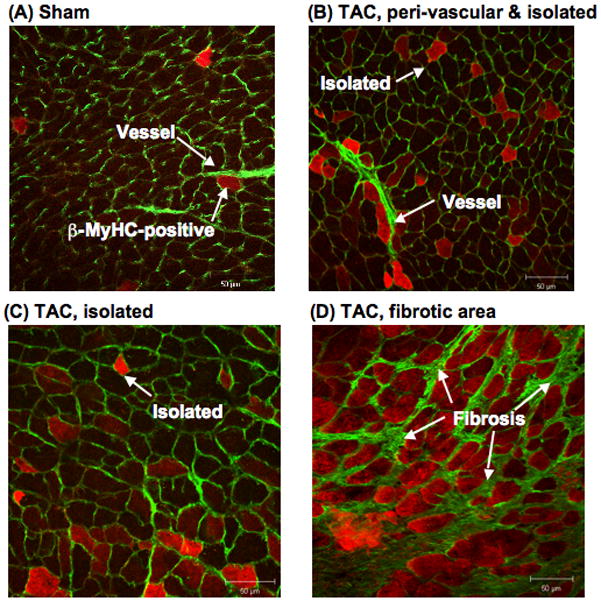
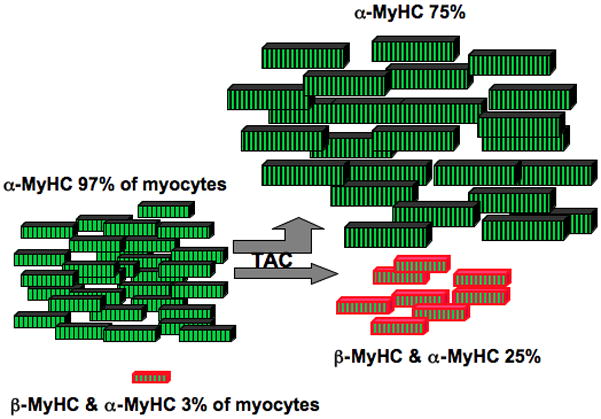
Methods and results: Myocytes were isolated from the left ventricle of male C57BL/6J mice after transverse aortic constriction (TAC), and the fraction of cells expressing endogenous β-MyHC was quantified by flow cytometry on 10,000 to 20,000 myocytes with use of a validated β-MyHC antibody. Side scatter by flow cytometry in the same cells was validated as an index of myocyte size. β-MyHC-positive myocytes constituted 3 ± 1% of myocytes in control hearts (n=12), increasing to 25 ± 10% at 3 days to 6 weeks after TAC (n=24, P<0.01). β-MyHC-positive myocytes did not enlarge with TAC and were smaller at all times than myocytes without β-MyHC (≈70% as large, P<0.001). β-MyHC-positive myocytes arose by addition of β-MyHC to α-MyHC and had more total MyHC after TAC than did the hypertrophied myocytes that had α-MyHC only. Myocytes positive for β-MyHC were found in discrete regions of the left ventricle in 3 patterns: perivascular, in areas with fibrosis, and in apparently normal myocardium.
Conclusions: β-MyHC protein is induced by pressure overload in a minor subpopulation of smaller cardiac myocytes. The hypertrophied myocytes after TAC have α-MyHC only. These data challenge the current paradigm of the fetal hypertrophic gene program and identify a new subpopulation of smaller working ventricular myocytes with more myosin.
Figures
References
-
- Lompre AM, Schwartz K, d’Albis A, Lacombe G, Van Thiem N, Swynghedauw B. Myosin isoenzyme redistribution in chronic heart overload. Nature. 1979;282:105–107. - PubMed
-
- Lompre AM, Nadal-Ginard B, Mahdavi V. Expression of the cardiac ventricular alpha- and beta-myosin heavy chain genes is developmentally and hormonally regulated. J Biol Chem. 1984;259:6437–6446. - PubMed
-
- Mercadier JJ, Lompre AM, Wisnewsky C, Samuel JL, Bercovici J, Swynghedauw B, Schwartz K. Myosin isoenzyme changes in several models of rat cardiac hypertrophy. Circ Res. 1981;49:525–532. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
