

A genome-wide approach to discovery of small RNAs involved in regulation of virulence in Vibrio cholerae
- PMID: 21779167
- PMCID: PMC3136459
- DOI: 10.1371/journal.ppat.1002126
A genome-wide approach to discovery of small RNAs involved in regulation of virulence in Vibrio cholerae
Abstract
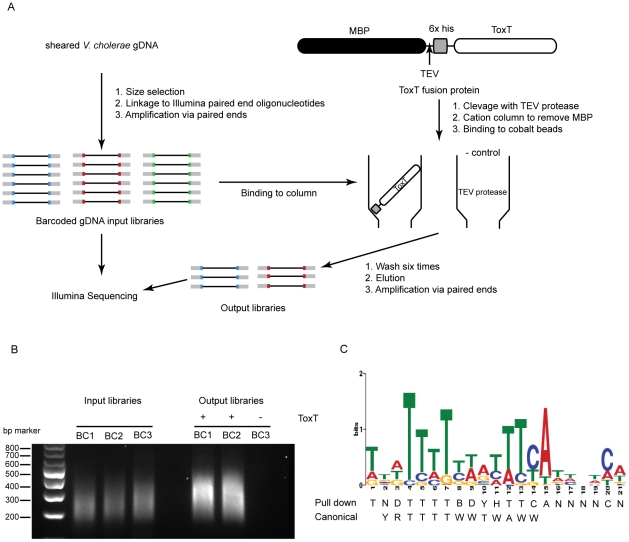
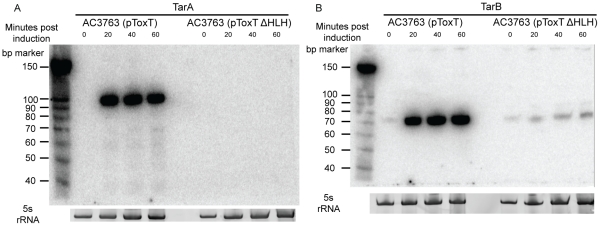
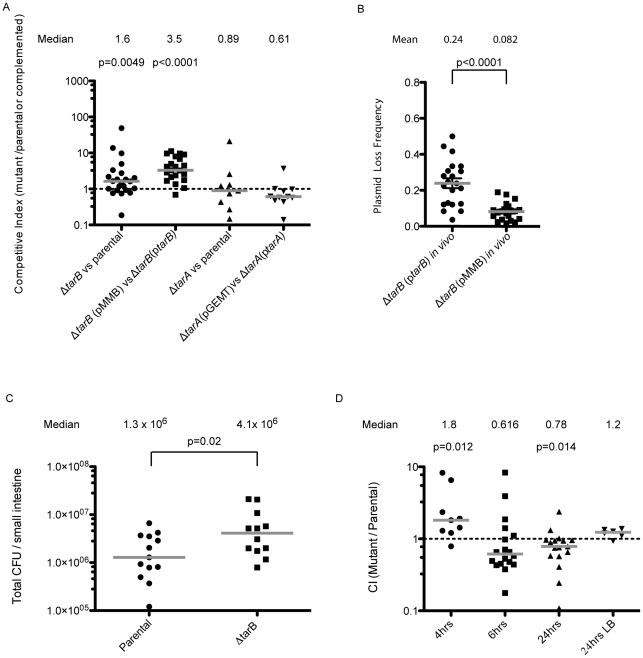
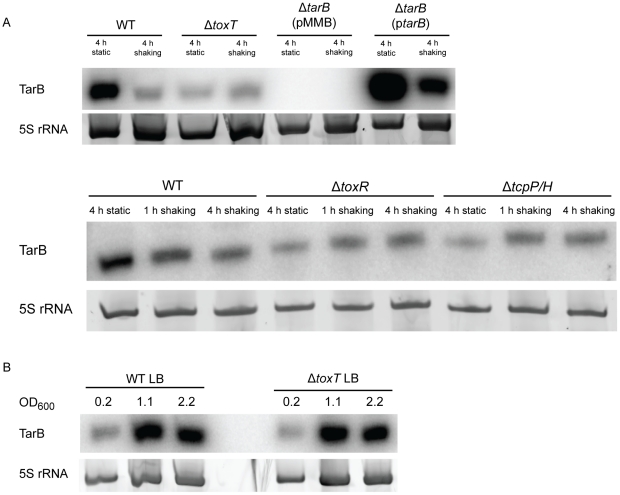
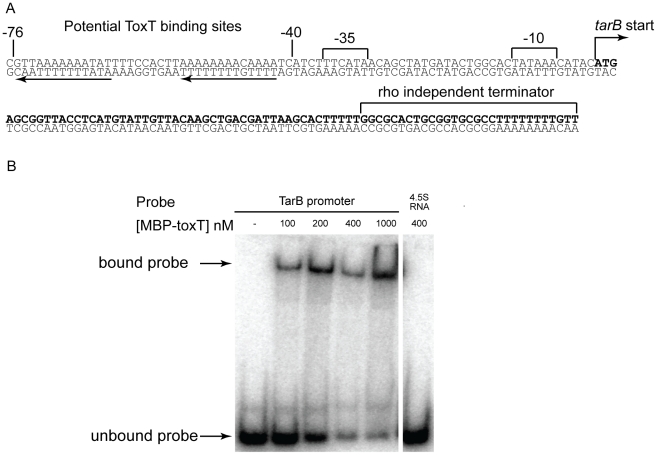
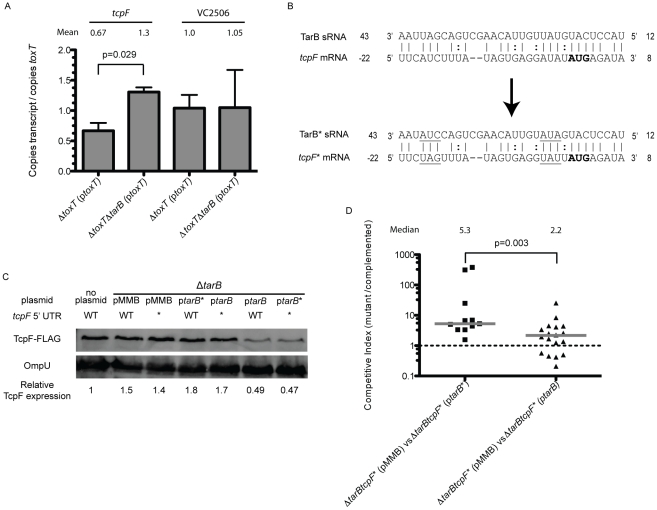
Small RNAs (sRNAs) are becoming increasingly recognized as important regulators in bacteria. To investigate the contribution of sRNA mediated regulation to virulence in Vibrio cholerae, we performed high throughput sequencing of cDNA generated from sRNA transcripts isolated from a strain ectopically expressing ToxT, the major transcriptional regulator within the virulence gene regulon. We compared this data set with ToxT binding sites determined by pulldown and deep sequencing to identify sRNA promoters directly controlled by ToxT. Analysis of the resulting transcripts with ToxT binding sites in cis revealed two sRNAs within the Vibrio Pathogenicity Island. When deletions of these sRNAs were made and the resulting strains were competed against the parental strain in the infant mouse model of V. cholerae colonization, one, TarB, displayed a variable colonization phenotype dependent on its physiological state at the time of inoculation. We identified a target of TarB as the mRNA for the secreted colonization factor, TcpF. We verified negative regulation of TcpF expression by TarB and, using point mutations that disrupted interaction between TarB and tpcF mRNA, showed that loss of this negative regulation was primarily responsible for the colonization phenotype observed in the TarB deletion mutant.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
