

The RpiR-like repressor IolR regulates inositol catabolism in Sinorhizobium meliloti
- PMID: 21784930
- PMCID: PMC3187398
- DOI: 10.1128/JB.05371-11
The RpiR-like repressor IolR regulates inositol catabolism in Sinorhizobium meliloti
Abstract
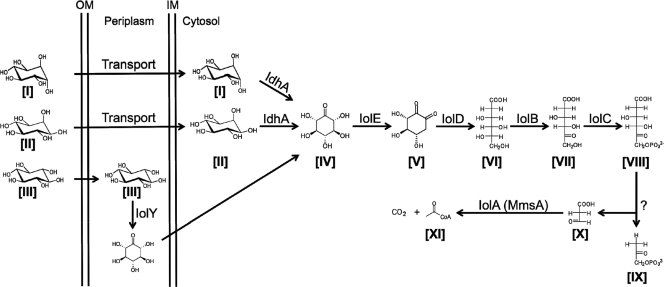
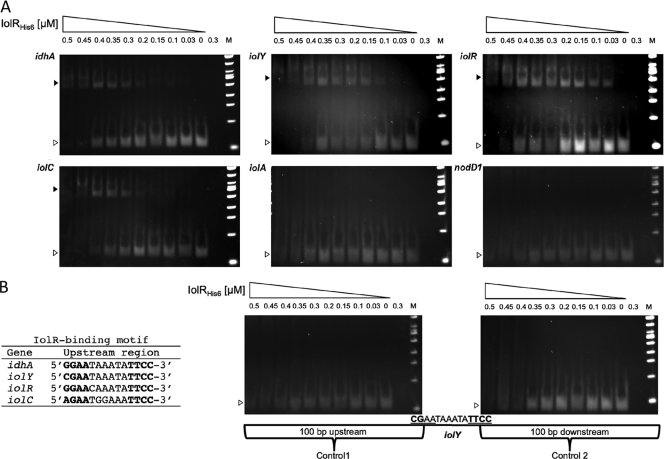
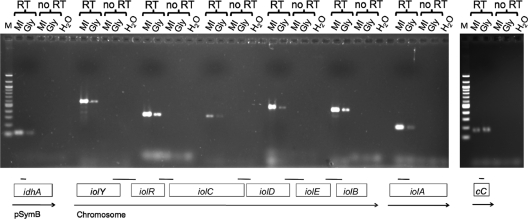
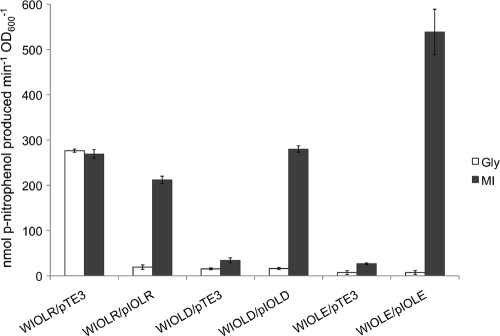
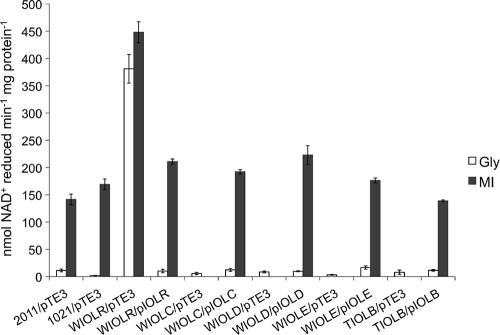
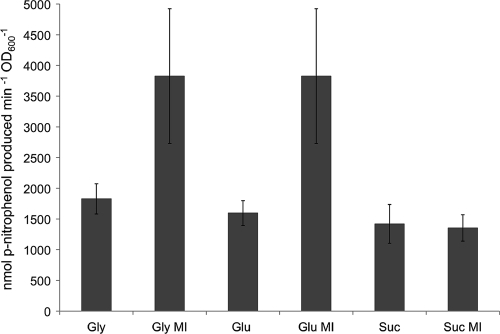
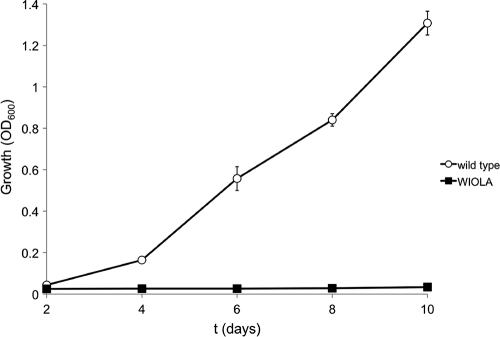
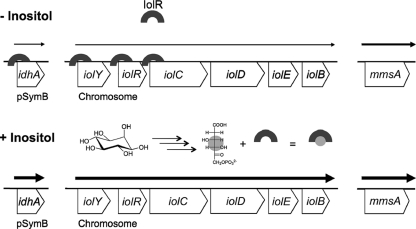
Sinorhizobium meliloti, the nitrogen-fixing symbiont of alfalfa, has the ability to catabolize myo-, scyllo-, and D-chiro-inositol. Functional inositol catabolism (iol) genes are required for growth on these inositol isomers, and they play a role during plant-bacterium interactions. The inositol catabolism genes comprise the chromosomally encoded iolA (mmsA) and the iolY(smc01163)RCDEB genes, as well as the idhA gene located on the pSymB plasmid. Reverse transcriptase assays showed that the iolYRCDEB genes are transcribed as one operon. The iol genes were weakly expressed without induction, but their expression was strongly induced by myo-inositol. The putative transcriptional regulator of the iol genes, IolR, belongs to the RpiR-like repressor family. Electrophoretic mobility shift assays demonstrated that IolR recognized a conserved palindromic sequence (5'-GGAA-N6-TTCC-3') in the upstream regions of the idhA, iolY, iolR, and iolC genes. Complementation assays found IolR to be required for the repression of its own gene and for the downregulation of the idhA-encoded myo-inositol dehydrogenase activity in the presence and absence of inositol. Further expression studies indicated that the late pathway intermediate 2-keto-5-deoxy-D-gluconic acid 6-phosphate (KDGP) functions as the true inducer of the iol genes. The iolA (mmsA) gene encoding methylmalonate semialdehyde dehydrogenase was not regulated by IolR. The S. meliloti iolA (mmsA) gene product seems to be involved in more than only the inositol catabolic pathway, since it was also found to be essential for valine catabolism, supporting its more recent annotation as mmsA.
Figures
References
-
- Bannerjee D., Sanders L. E., Sokatch J. R. 1970. Properties of purified methylmalonate semialdehyde dehydrogenase of Pseudomonas aeruginosa. J. Biol. Chem. 245:1828–1835 - PubMed
-
- Beringer J. E. 1974. R factor transfer in Rhizobium leguminosarum. J. Gen. Microbiol. 84:188–198 - PubMed
-
- Berman T., Magasanik B. 1966. The pathway of myo-inositol degradation in Aerobacter aerogenes: dehydrogenation and dehydratation. J. Biol. Chem. 241:800–806 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
