

Tapping natural reservoirs of homing endonucleases for targeted gene modification
- PMID: 21784983
- PMCID: PMC3156218
- DOI: 10.1073/pnas.1107719108
Tapping natural reservoirs of homing endonucleases for targeted gene modification
Abstract
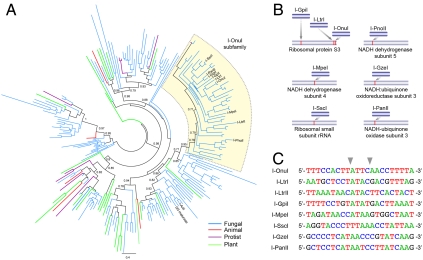
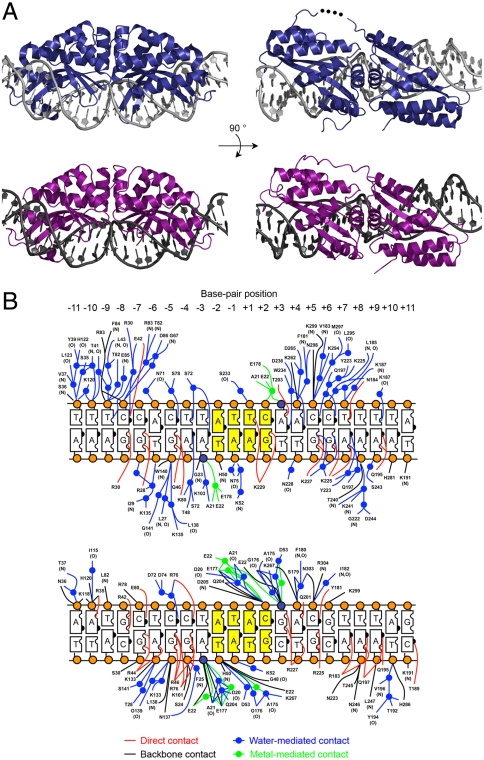
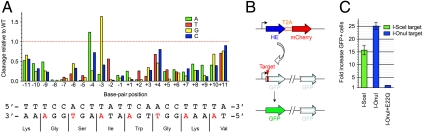
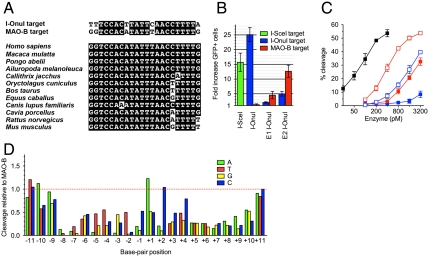
Homing endonucleases mobilize their own genes by generating double-strand breaks at individual target sites within potential host DNA. Because of their high specificity, these proteins are used for "genome editing" in higher eukaryotes. However, alteration of homing endonuclease specificity is quite challenging. Here we describe the identification and phylogenetic analysis of over 200 naturally occurring LAGLIDADG homing endonucleases (LHEs). Biochemical and structural characterization of endonucleases from one clade within the phylogenetic tree demonstrates strong conservation of protein structure contrasted against highly diverged DNA target sites and indicates that a significant fraction of these proteins are sufficiently stable and active to serve as engineering scaffolds. This information was exploited to create a targeting enzyme to disrupt the endogenous monoamine oxidase B gene in human cells. The ubiquitous presence and diversity of LHEs described in this study may facilitate the creation of many tailored nucleases for genome editing.
Conflict of interest statement
Conflict of interest statement: B.L.S. and A.M.S. are founders of a biotechnology company (Precision Genome Engineering) that conducts engineering and selection studies on homing endonucleases for gene targeting.
Figures
References
-
- Urnov FD, Rebar EJ, Holmes MC, Zhang HS, Gregory PD. Genome editing with engineered zinc finger nucleases. Nat Rev Genet. 2010;11:636–646. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
