

Cytoskeletal polarity mediates localized induction of the heart progenitor lineage
- PMID: 21785423
- PMCID: PMC3149722
- DOI: 10.1038/ncb2291
Cytoskeletal polarity mediates localized induction of the heart progenitor lineage
Abstract
Cells must make appropriate fate decisions within a complex and dynamic environment. In vitro studies indicate that the cytoskeleton acts as an integrative platform for this environmental input. External signals regulate cytoskeletal dynamics and the cytoskeleton reciprocally modulates signal transduction. However, in vivo studies linking cytoskeleton/signalling interactions to embryonic cell fate specification remain limited. Here we show that the cytoskeleton modulates heart progenitor cell fate. Our studies focus on differential induction of heart fate in the basal chordate Ciona intestinalis. We have found that differential induction does not simply reflect differential exposure to the inductive signal. Instead, pre-cardiac cells employ polarized, invasive protrusions to localize their response to an ungraded signal. Through targeted manipulation of the cytoskeletal regulator CDC42, we are able to depolarize protrusive activity and generate uniform heart progenitor fate specification. Furthermore, we are able to restore differential induction by repolarizing protrusive activity. These findings illustrate how bi-directional interactions between intercellular signalling and the cytoskeleton can influence embryonic development. In particular, these studies highlight the potential for dynamic cytoskeletal changes to refine cell fate specification in response to crude signal gradients.
Figures
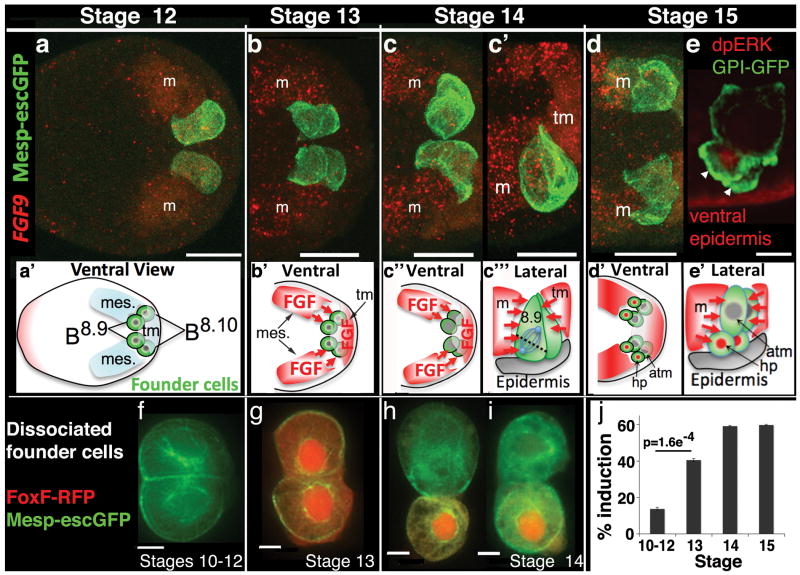
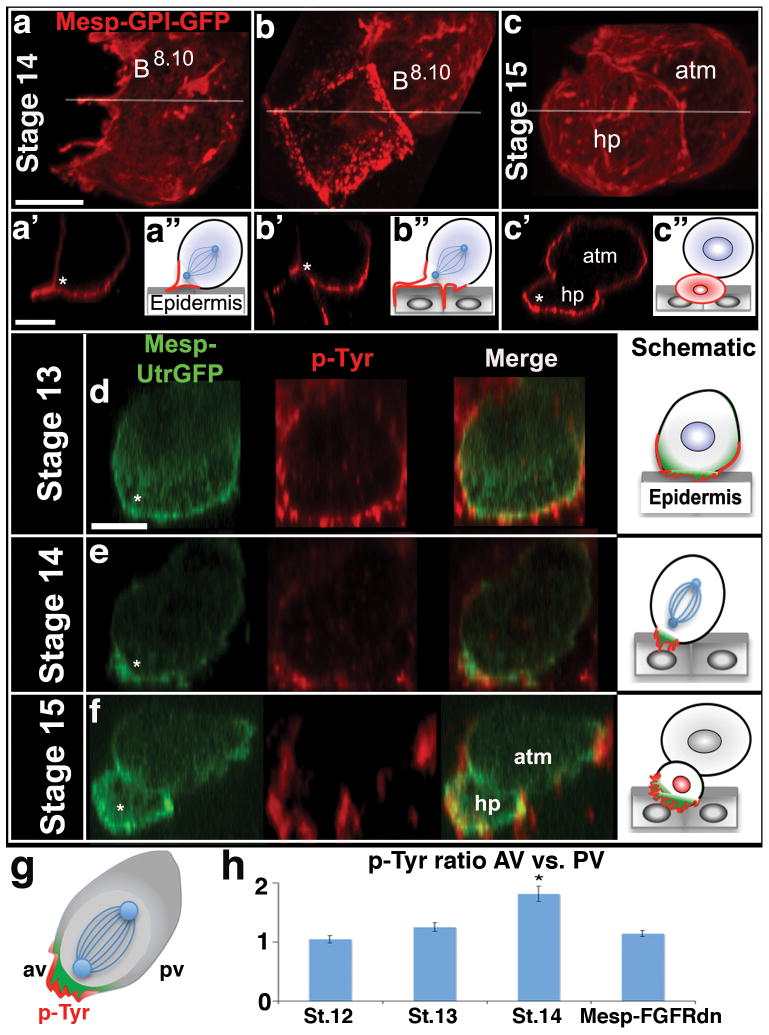
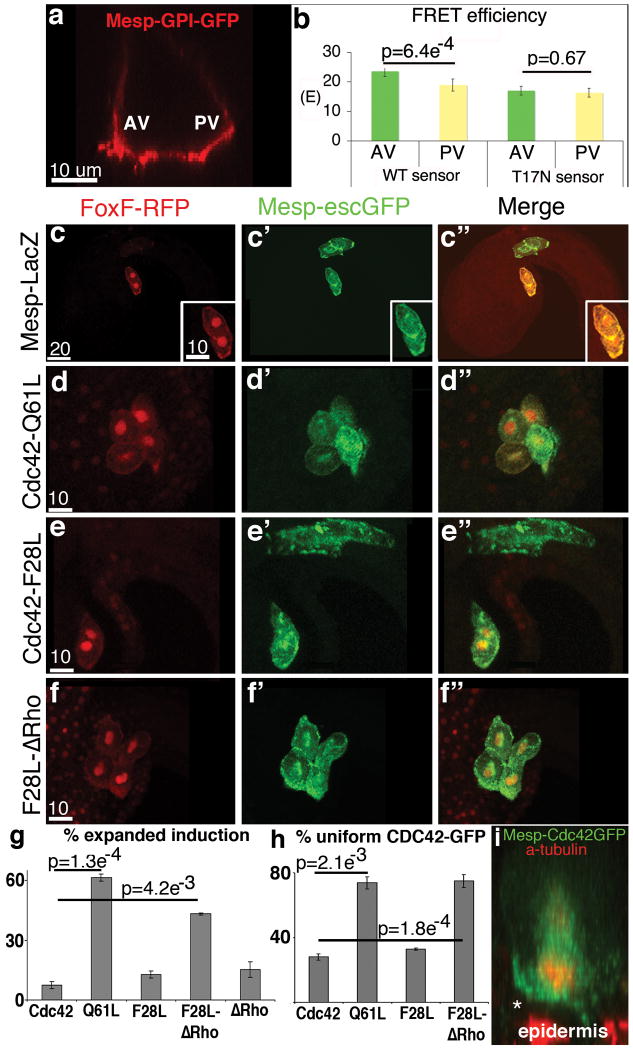
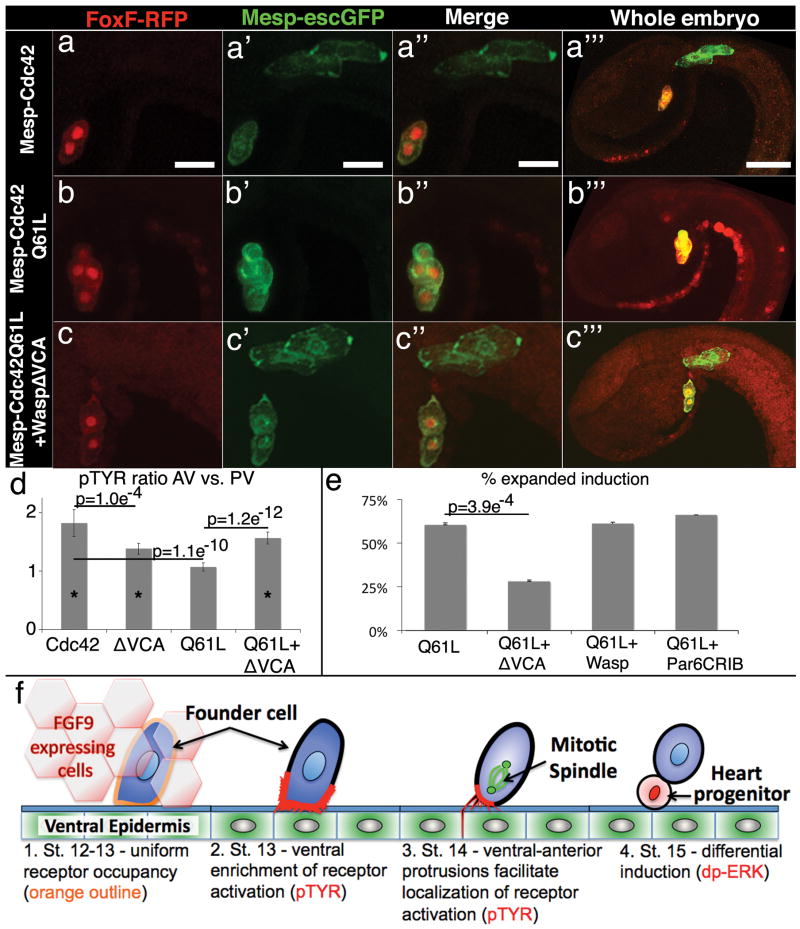
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
