

Pattern separation in the hippocampus
- PMID: 21788086
- PMCID: PMC3183227
- DOI: 10.1016/j.tins.2011.06.006
Pattern separation in the hippocampus
Abstract
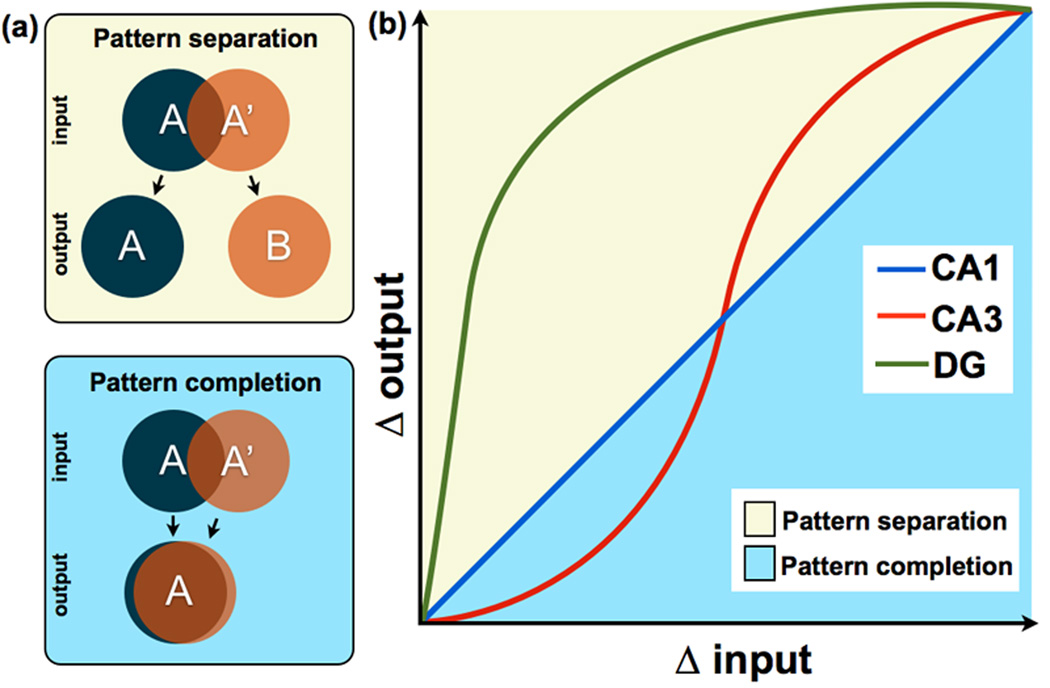
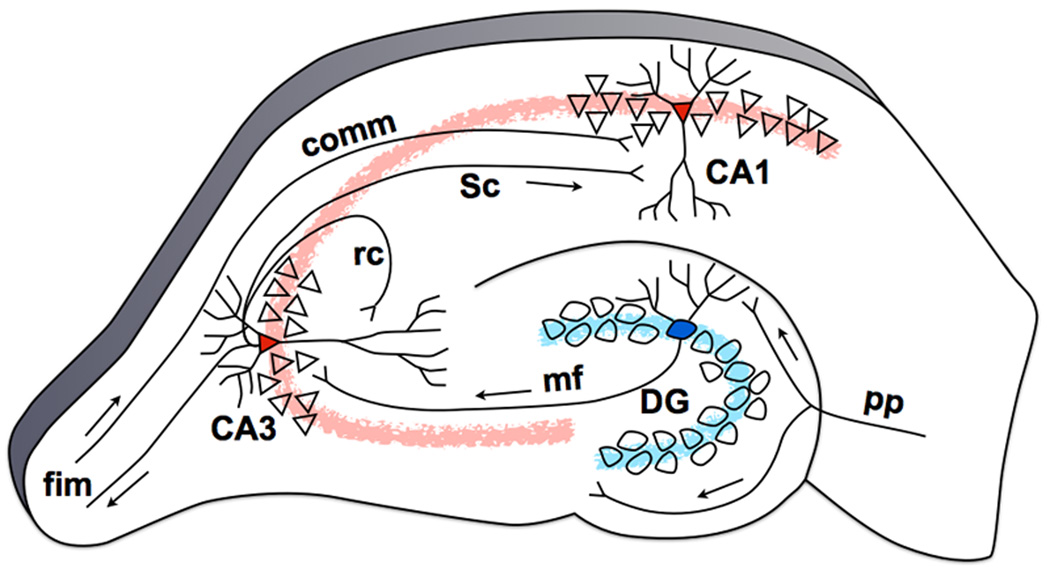
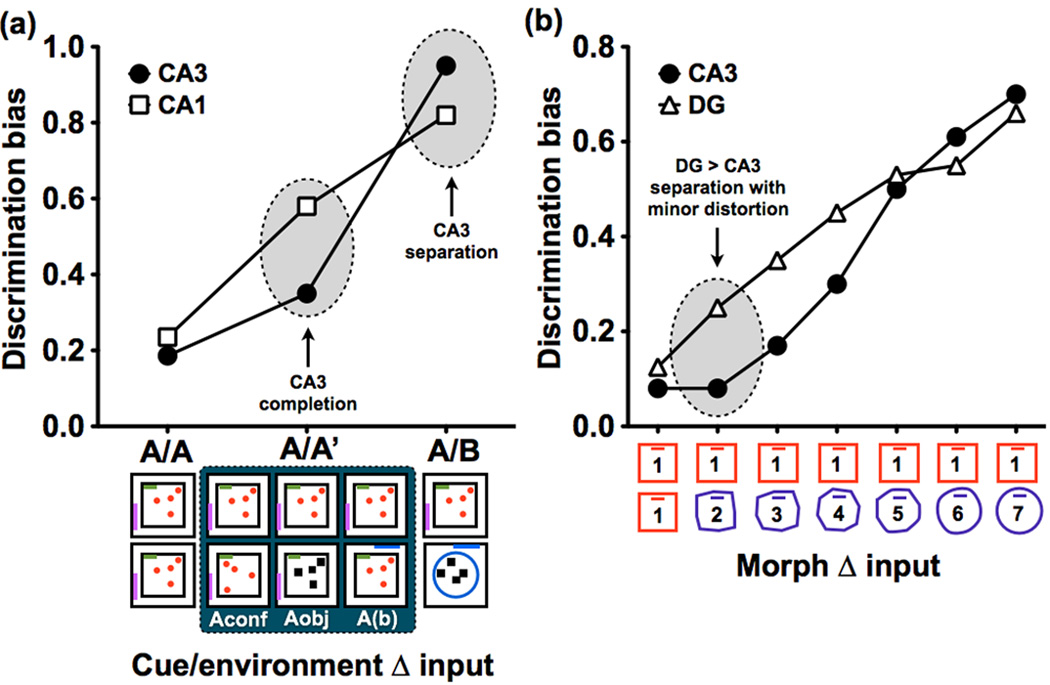
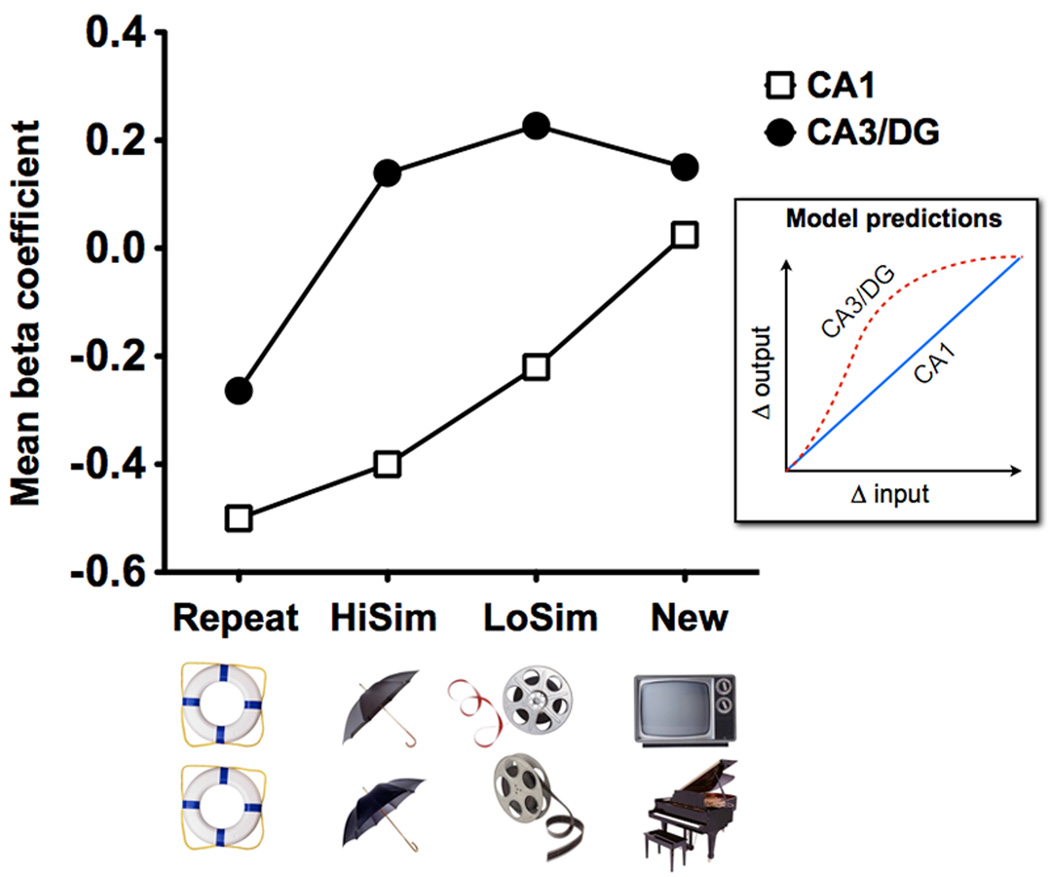
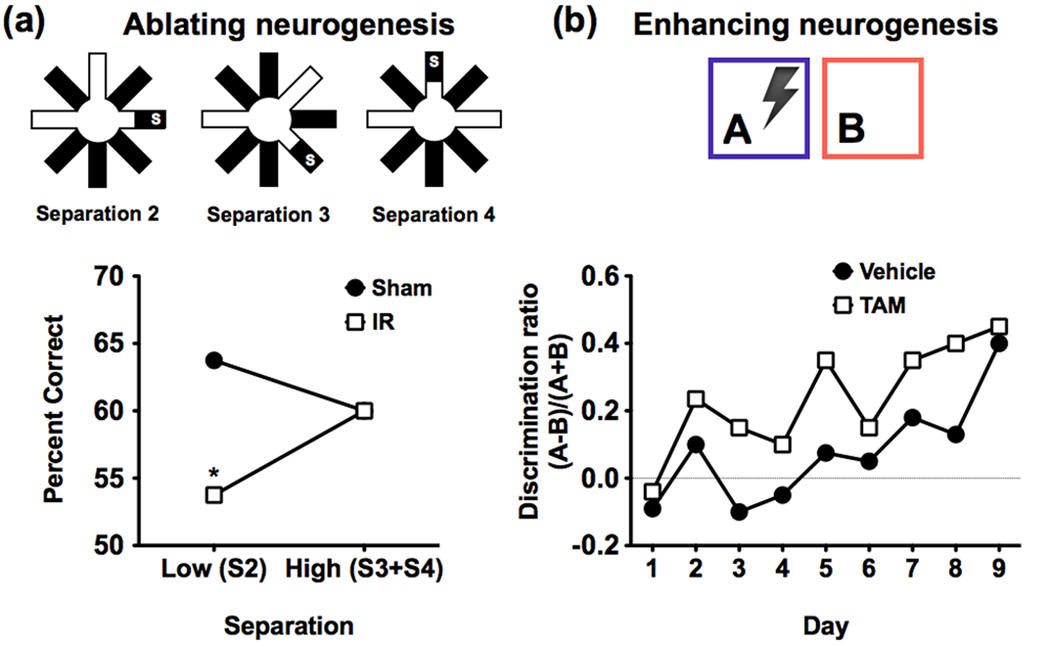
The ability to discriminate among similar experiences is a crucial feature of episodic memory. This ability has long been hypothesized to require the hippocampus, and computational models suggest that it is dependent on pattern separation. However, empirical data for the role of the hippocampus in pattern separation have not been available until recently. This review summarizes data from electrophysiological recordings, lesion studies, immediate-early gene imaging, transgenic mouse models, as well as human functional neuroimaging, that provide convergent evidence for the involvement of particular hippocampal subfields in this key process. We discuss the impact of aging and adult neurogenesis on pattern separation, and also highlight several challenges to linking across species and approaches, and suggest future directions for investigation.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
References
-
- Marr D. Simple memory: a theory for archicortex. Philos Trans R Soc Lond B Biol Sci. 1971;262:23–81. - PubMed
-
- McClelland JL, et al. Why there are complementary learning systems in the hippocampus and neocortex: insights from the successes and failures of connectionist models of learning and memory. Psychol Rev. 1995;102:419–457. - PubMed
-
- Norman KA, O'Reilly RC. Modeling hippocampal and neocortical contributions to recognition memory: a complementary-learning-systems approach. Psychol Rev. 2003;110:611–646. - PubMed
-
- McNaughton BL, Morris RG. Hippocampal synaptic enhancement and information storage within a distributed memory system. Trends Neurosci. 1987;10:408–415.
-
- Shapiro ML, Olton DS. Hippocampal function and interference. In: Schacter DL, Tulving E, editors. Memory Systems 1994. MIT Press; 1994. pp. 141–146.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
