

Endothelial cells create a stem cell niche in glioblastoma by providing NOTCH ligands that nurture self-renewal of cancer stem-like cells
- PMID: 21788346
- PMCID: PMC3355476
- DOI: 10.1158/0008-5472.CAN-10-4269
Endothelial cells create a stem cell niche in glioblastoma by providing NOTCH ligands that nurture self-renewal of cancer stem-like cells
Abstract
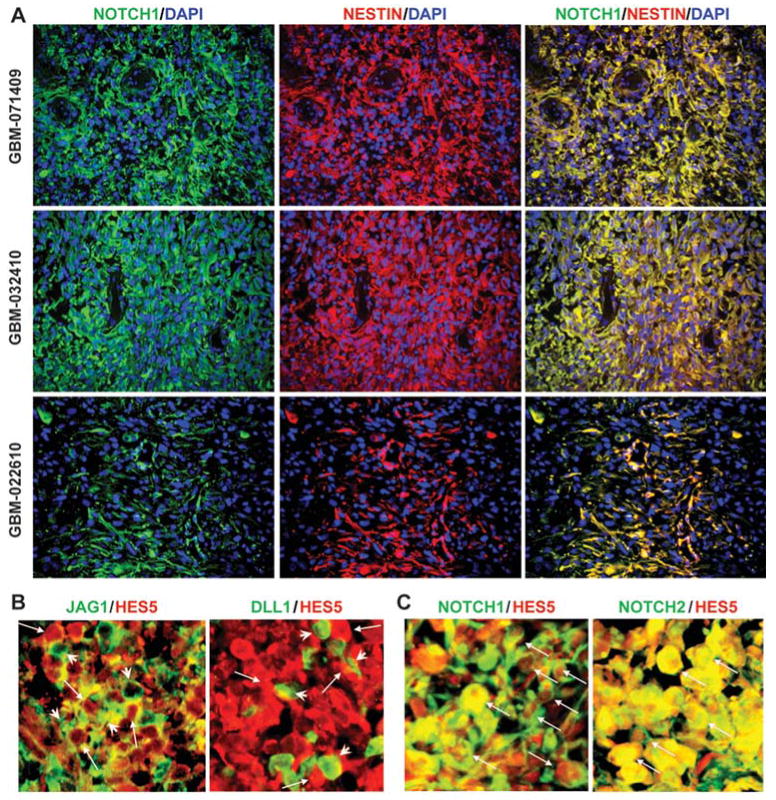
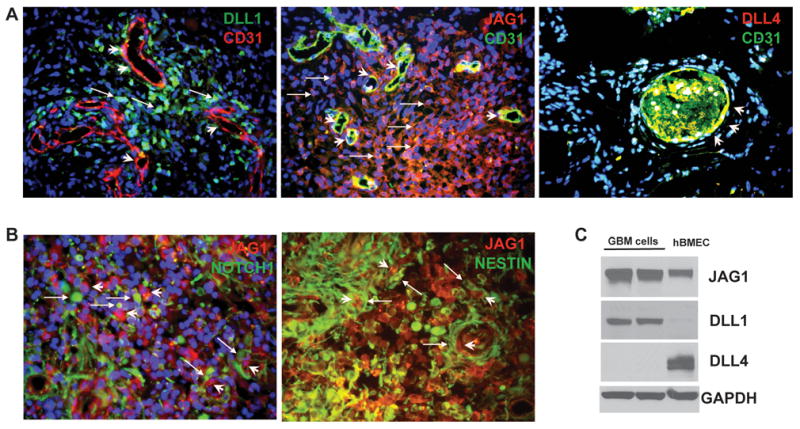
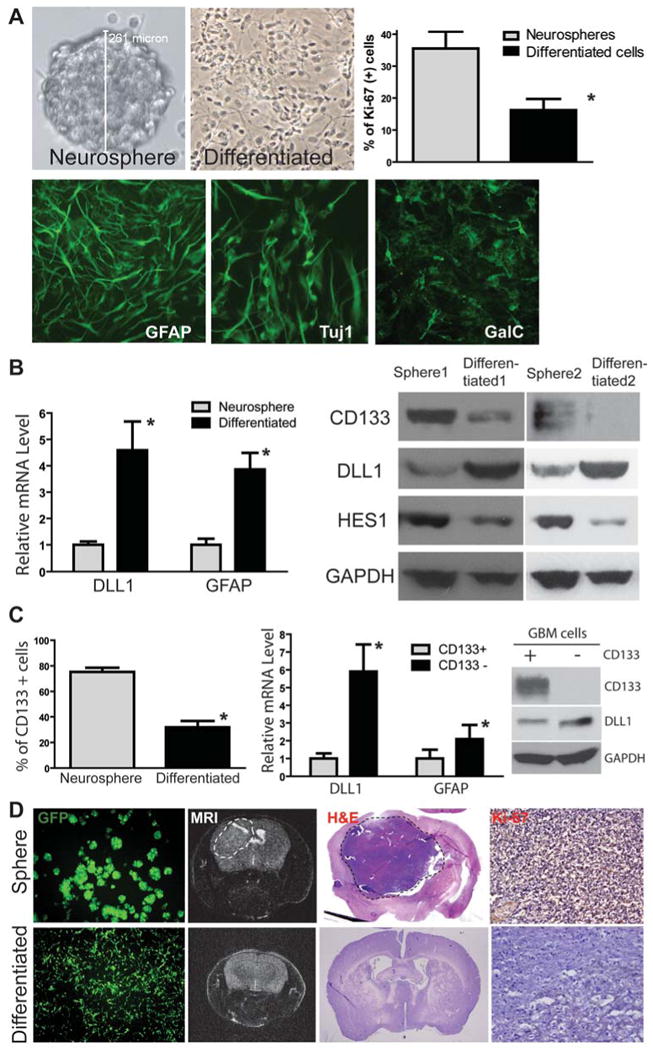
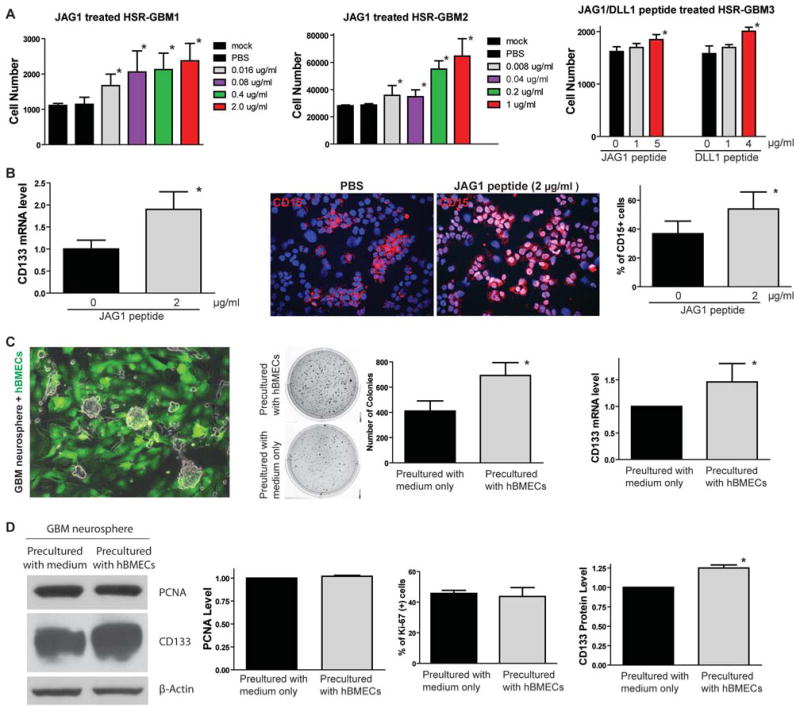
One important function of endothelial cells in glioblastoma multiforme (GBM) is to create a niche that helps promote self-renewal of cancer stem-like cells (CSLC). However, the underlying molecular mechanism for this endothelial function is not known. Since activation of NOTCH signaling has been found to be required for propagation of GBM CSLCs, we hypothesized that the GBM endothelium may provide the source of NOTCH ligands. Here, we report a corroboration of this concept with a demonstration that NOTCH ligands are expressed in endothelial cells adjacent to NESTIN and NOTCH receptor-positive cancer cells in primary GBMs. Coculturing human brain microvascular endothelial cells (hBMEC) or NOTCH ligand with GBM neurospheres promoted GBM cell growth and increased CSLC self-renewal. Notably, RNAi-mediated knockdown of NOTCH ligands in hBMECs abrogated their ability to induce CSLC self-renewal and GBM tumor growth, both in vitro and in vivo. Thus, our findings establish that NOTCH activation in GBM CSLCs is driven by juxtacrine signaling between tumor cells and their surrounding endothelial cells in the tumor microenvironment, suggesting that targeting both CSLCs and their niche may provide a novel strategy to deplete CSLCs and improve GBM treatment.
Figures
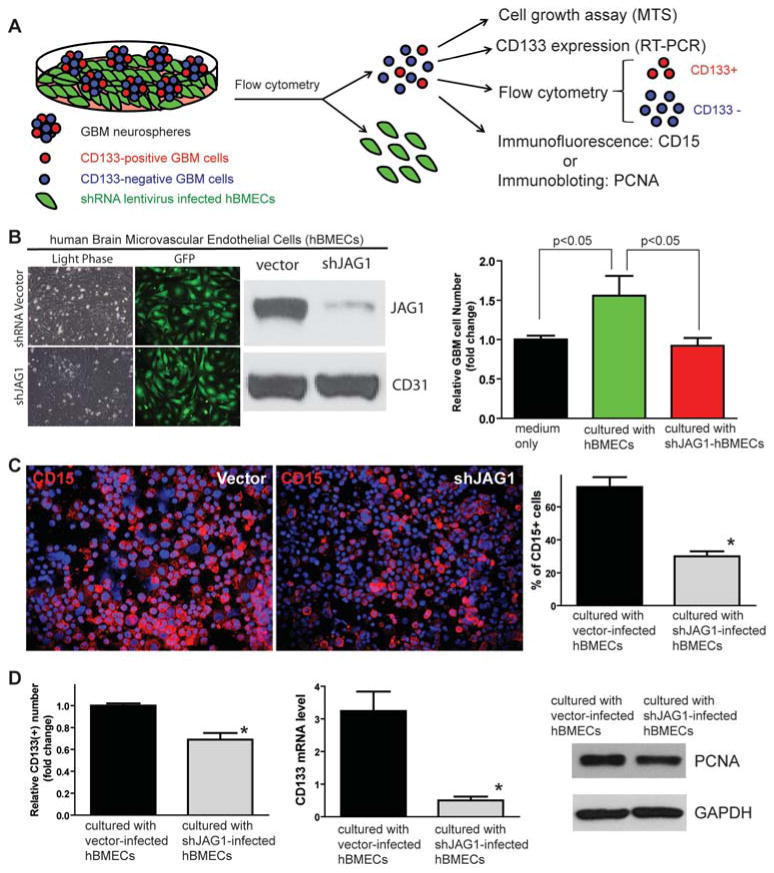
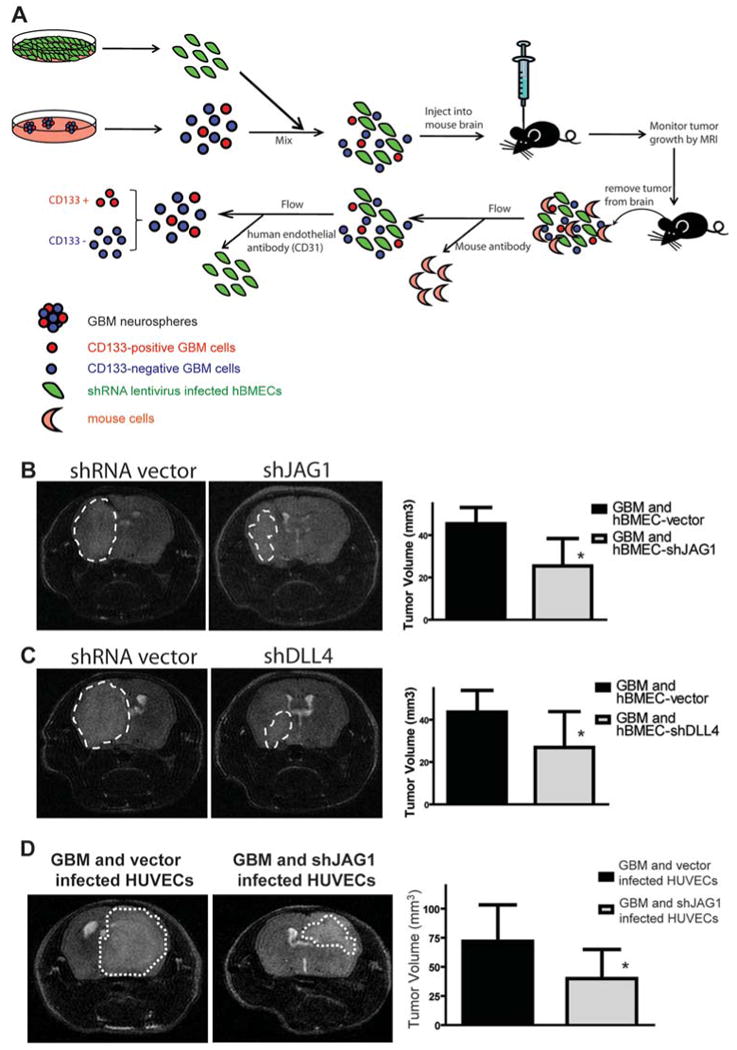
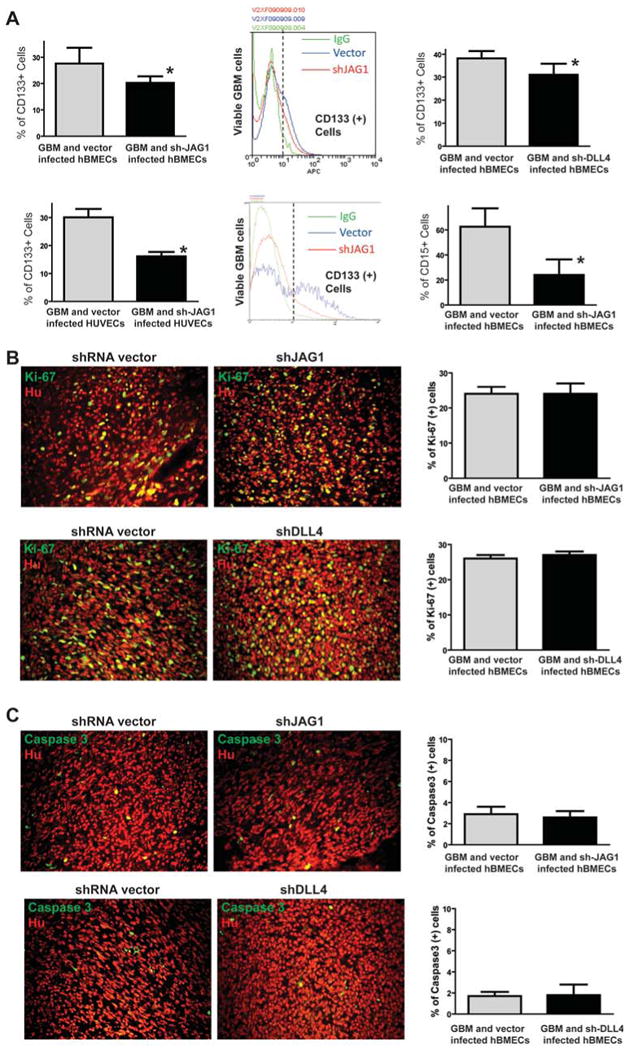
References
-
- Reardon DA, Rich JN, Friedman HS, Bigner DD. Recent advances in the treatment of malignant astrocytoma. J Clin Oncol. 2006 Mar 10;24(8):1253–65. - PubMed
-
- Stupp R, Mason WP, van den Bent MJ, Weller M, Fisher B, Taphoorn MJ, et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med. 2005 Mar 10;352(10):987–96. - PubMed
-
- Reya T, Morrison SJ, Clarke MF, Weissman IL. Stem cells, cancer, and cancer stem cells. Nature. 2001 Nov 1;414(6859):105–11. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
