

Analysis of a range of catabolic mutants provides evidence that phytanoyl-coenzyme A does not act as a substrate of the electron-transfer flavoprotein/electron-transfer flavoprotein:ubiquinone oxidoreductase complex in Arabidopsis during dark-induced senescence
- PMID: 21788362
- PMCID: PMC3221279
- DOI: 10.1104/pp.111.182188
Analysis of a range of catabolic mutants provides evidence that phytanoyl-coenzyme A does not act as a substrate of the electron-transfer flavoprotein/electron-transfer flavoprotein:ubiquinone oxidoreductase complex in Arabidopsis during dark-induced senescence
Abstract
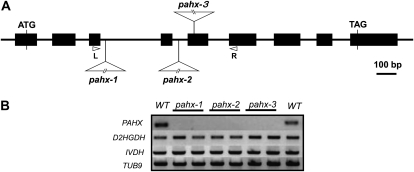
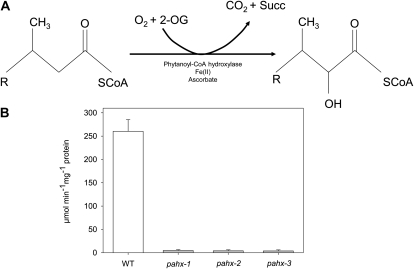
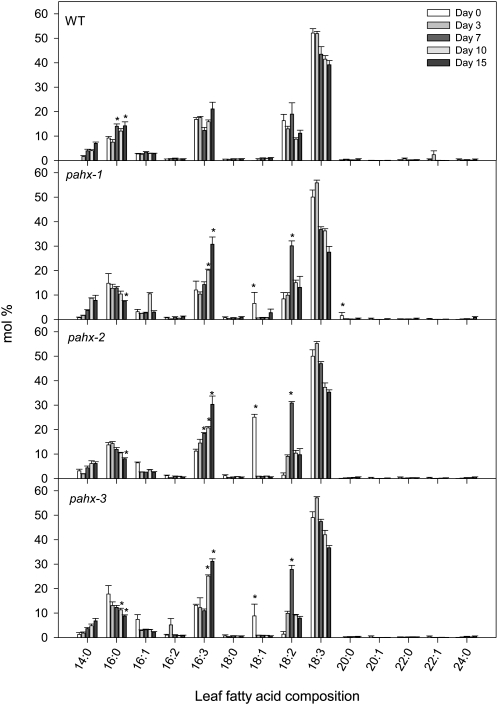
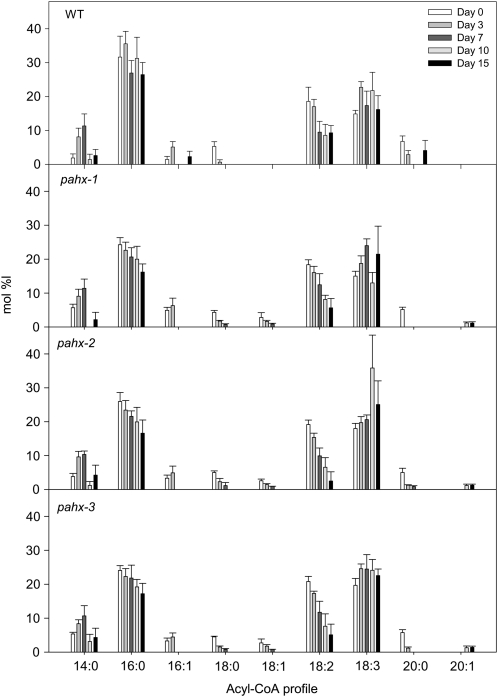
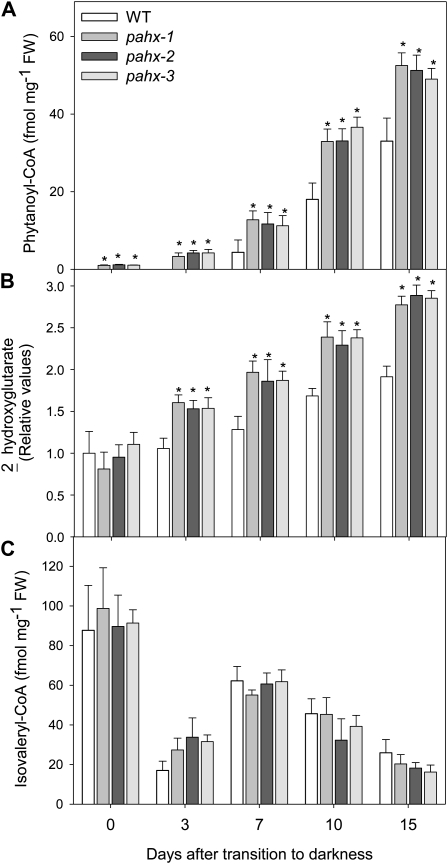
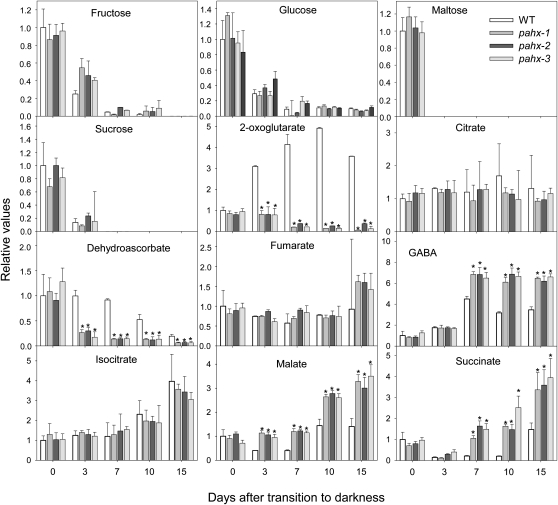
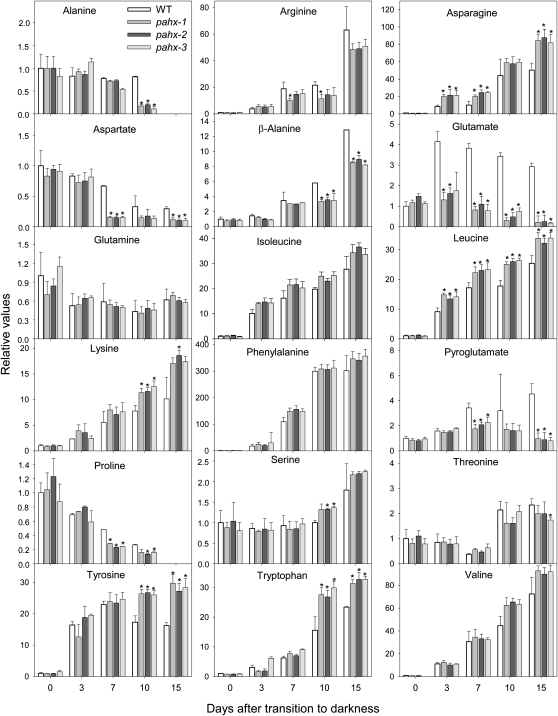
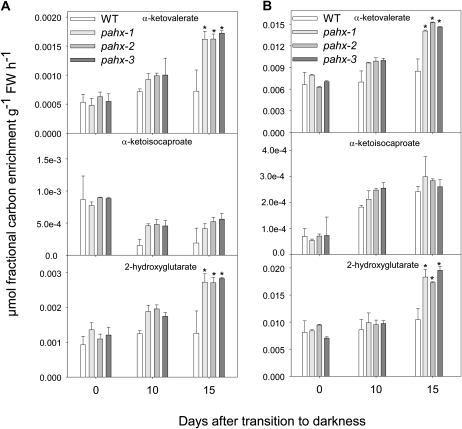
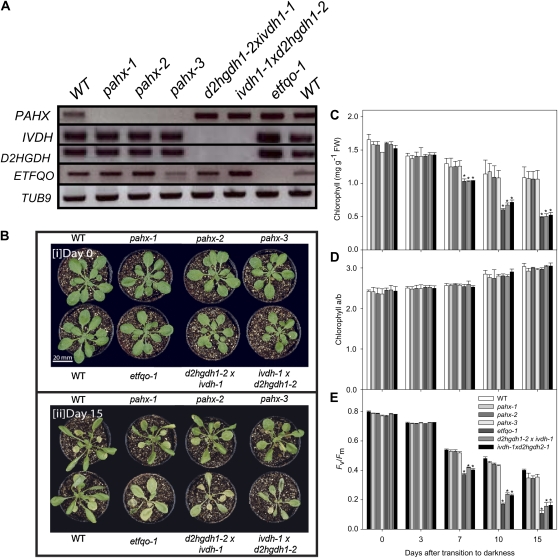
The process of dark-induced senescence in plants is not fully understood, however, the functional involvement of an electron-transfer flavoprotein/electron-transfer flavoprotein:ubiquinone oxidoreductase (ETF/ETFQO), has been demonstrated. Recent studies have revealed that the enzymes isovaleryl-coenzyme A (CoA) dehydrogenase and 2-hydroxyglutarate dehydrogenase act as important electron donors to this complex. In addition both enzymes play a role in the breakdown of cellular carbon storage reserves with isovaleryl-CoA dehydrogenase being involved in degradation of the branched-chain amino acids, phytol, and lysine while 2-hydroxyglutarate dehydrogenase is exclusively involved in lysine degradation. Given that the chlorophyll breakdown intermediate phytanoyl-CoA accumulates dramatically both in knockout mutants of the ETF/ETFQO complex and of isovaleryl-CoA dehydrogenase following growth in extended dark periods we have investigated the direct importance of chlorophyll breakdown for the supply of carbon and electrons during this process. For this purpose we isolated three independent Arabidopsis (Arabidopsis thaliana) knockout mutants of phytanoyl-CoA 2-hydroxylase and grew them under the same extended darkness regime as previously used. Despite the fact that these mutants accumulated phytanoyl-CoA and also 2-hydroxyglutarate they exhibited no morphological changes in comparison to the other mutants previously characterized. These results are consistent with a single entry point of phytol breakdown into the ETF/ETFQO system and furthermore suggest that phytol is not primarily metabolized by this pathway. Furthermore analysis of isovaleryl-CoA dehydrogenase/2-hydroxyglutarate dehydrogenase double mutants generated here suggest that these two enzymes essentially account for the entire electron input via the ETF complex.
Figures
References
-
- Araújo WL, Ishizaki K, Nunes-Nesi A, Larson TR, Tohge T, Krahnert I, Witt S, Obata T, Schauer N, Graham IA, et al. (2010) Identification of the 2-hydroxyglutarate and isovaleryl-CoA dehydrogenases as alternative electron donors linking lysine catabolism to the electron transport chain of Arabidopsis mitochondria. Plant Cell 22: 1549–1563 - PMC - PubMed
-
- Araújo WL, Tohge T, Ishizaki K, Leaver CJ, Fernie AR. (2011) Protein degradation—an alternative respiratory substrate for stressed plants. Trends Plant Sci (in press) - PubMed
-
- Beckmann JD, Frerman FE. (1985) Electron-transfer flavoprotein-ubiquinone oxidoreductase from pig liver: purification and molecular, redox, and catalytic properties. Biochemistry 24: 3913–3921 - PubMed
-
- Bradford MM. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248–254 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
