

Gene targeting implicates Cdc42 GTPase in GPVI and non-GPVI mediated platelet filopodia formation, secretion and aggregation
- PMID: 21789221
- PMCID: PMC3138762
- DOI: 10.1371/journal.pone.0022117
Gene targeting implicates Cdc42 GTPase in GPVI and non-GPVI mediated platelet filopodia formation, secretion and aggregation
Abstract
Background: Cdc42 and Rac1, members of the Rho family of small GTPases, play critical roles in actin cytoskeleton regulation. We have shown previously that Rac1 is involved in regulation of platelet secretion and aggregation. However, the role of Cdc42 in platelet activation remains controversial. This study was undertaken to better understand the role of Cdc42 in platelet activation.
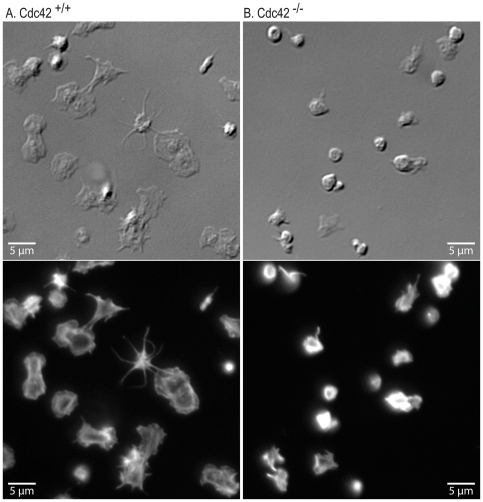
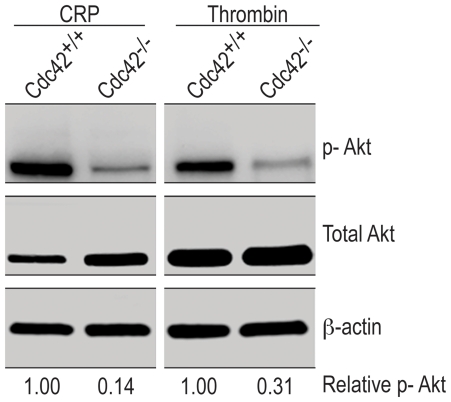
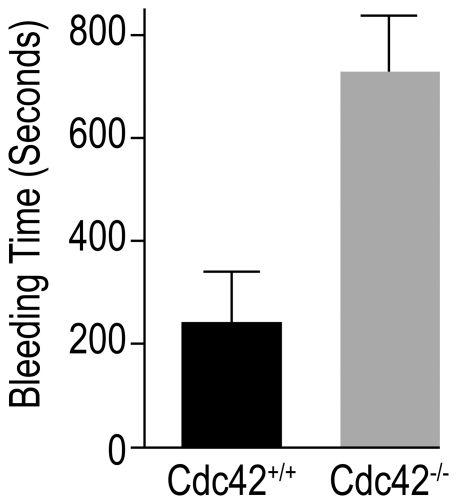
Methodology/principal findings: We utilized the Mx-cre;Cdc42(lox/lox) inducible mice with transient Cdc42 deletion to investigate the involvement of Cdc42 in platelet function. The Cdc42-deficient mice exhibited a significantly reduced platelet count than the matching Cdc42(+/+) mice. Platelets isolated from Cdc42(-/-), as compared to Cdc42(+/+), mice exhibited (a) diminished phosphorylation of PAK1/2, an effector molecule of Cdc42, (b) inhibition of filopodia formation on immobilized CRP or fibrinogen, (c) inhibition of CRP- or thrombin-induced secretion of ATP and release of P-selectin, (d) inhibition of CRP, collagen or thrombin induced platelet aggregation, and (e) minimal phosphorylation of Akt upon stimulation with CRP or thrombin. The bleeding times were significantly prolonged in Cdc42(-/-) mice compared with Cdc42(+/+) mice.
Conclusion/significance: Our data demonstrate that Cdc42 is required for platelet filopodia formation, secretion and aggregation and therefore plays a critical role in platelet mediated hemostasis and thrombosis.
Conflict of interest statement
Figures





References
-
- Etienne-Manneville S, Hall A. Rho GTPases in cell biology. Nature. 2002;420:629–635. - PubMed
-
- Erickson JW, Cerione RA. Multiple roles for Cdc42 in cell regulation. Curr Opin Cell Biol. 2001;13:153–157. - PubMed
-
- Ridley AJ. Rho GTPases and actin dynamics in membrane protrusions and vesicle trafficking. Trends Cell Biol. 2006;16:522–529. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
