

An emerging role for TOR signaling in mammalian tissue and stem cell physiology
- PMID: 21791526
- PMCID: PMC3143559
- DOI: 10.1242/dev.058230
An emerging role for TOR signaling in mammalian tissue and stem cell physiology
Abstract
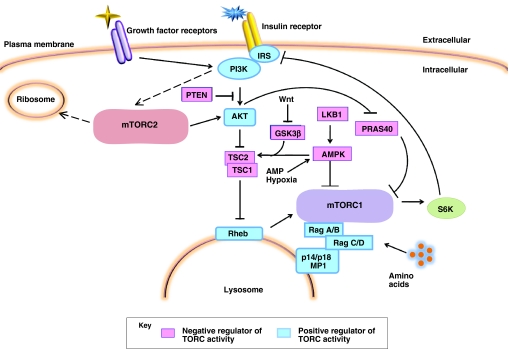
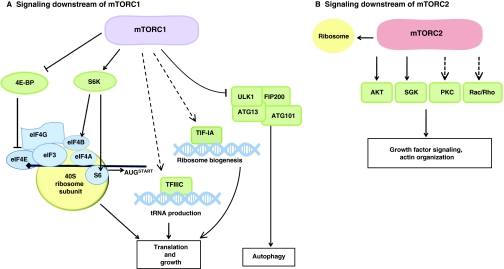
The mammalian target of rapamycin (mTOR) is a kinase that responds to a myriad of signals, ranging from nutrient availability and energy status, to cellular stressors, oxygen sensors and growth factors. The finely tuned response of mTOR to these stimuli results in alterations to cell metabolism and cell growth. Recent studies of conditional knockouts of mTOR pathway components in mice have affirmed the role of mTOR signaling in energy balance, both at the cell and whole organism levels. Such studies have also highlighted a role for mTOR in stem cell homeostasis and lifespan determination. Here, we discuss the molecular mechanisms of TOR signaling and review recent in vitro and in vivo studies of mTOR tissue-specific activities in mammals.
Figures
References
-
- Alessi D. R., Deak M., Casamayor A., Caudwell F. B., Morrice N., Norman D. G., Gaffney P., Reese C. B., MacDougall C. N., Harbison D., et al. (1997). 3-Phosphoinositide-dependent protein kinase-1 (PDK1): structural and functional homology with the Drosophila DSTPK61 kinase. Curr. Biol. 7, 776-789 - PubMed
-
- Artal-Sanz M., Tavernarakis N. (2009). Prohibitin couples diapause signalling to mitochondrial metabolism during ageing in C. elegans. Nature 461, 793-797 - PubMed
-
- Bateman J. M., McNeill H. (2004). Temporal control of differentiation by the insulin receptor/tor pathway in Drosophila. Cell 119, 87-96 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
