

Balanced interactions between Lyn, the p85alpha regulatory subunit of class I(A) phosphatidylinositol-3-kinase, and SHIP are essential for mast cell growth and maturation
- PMID: 21791602
- PMCID: PMC3187372
- DOI: 10.1128/MCB.05750-11
Balanced interactions between Lyn, the p85alpha regulatory subunit of class I(A) phosphatidylinositol-3-kinase, and SHIP are essential for mast cell growth and maturation
Abstract
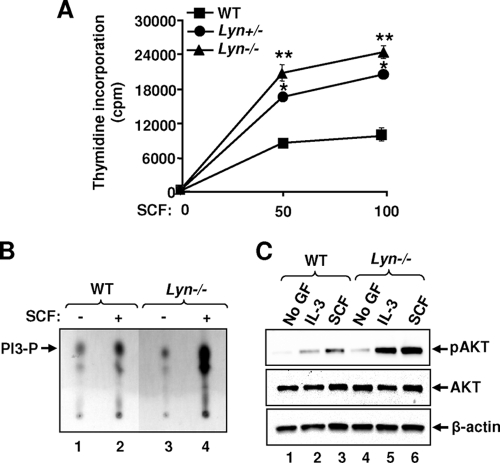
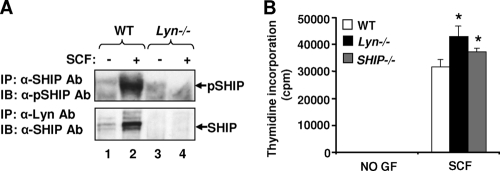
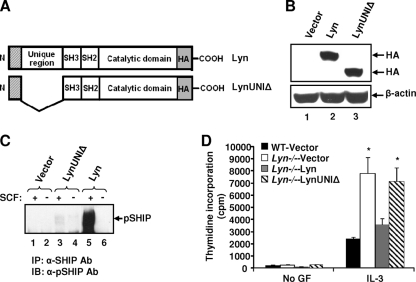
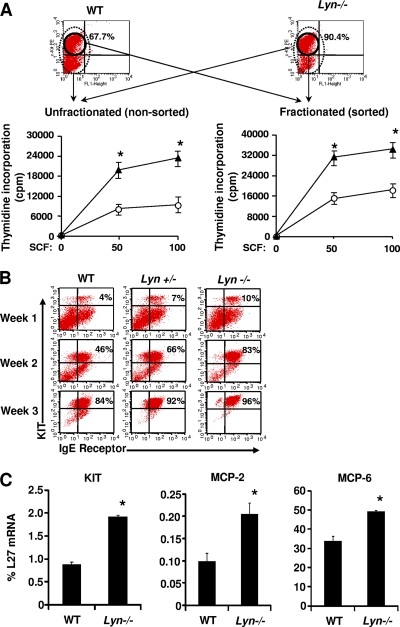
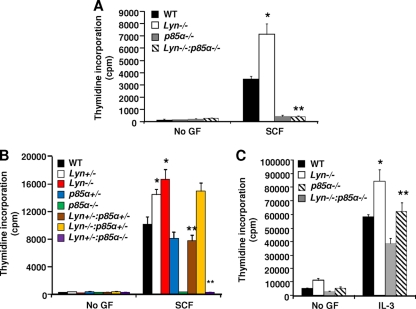
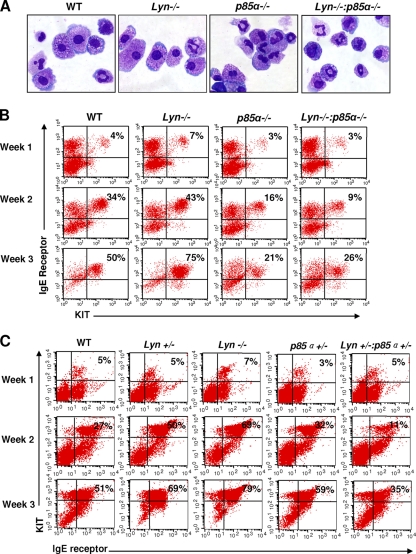
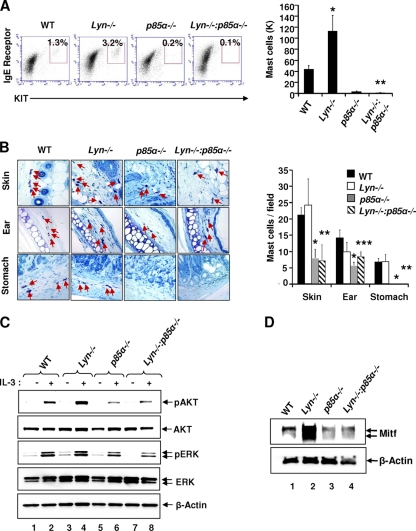
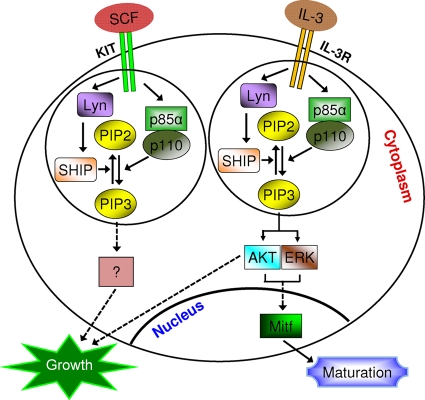
The growth and maturation of bone marrow-derived mast cells (BMMCs) from precursors are regulated by coordinated signals from multiple cytokine receptors, including KIT. While studies conducted using mutant forms of these receptors lacking the binding sites for Src family kinases (SFKs) and phosphatidylinositol-3-kinase (PI3K) suggest a role for these signaling molecules in regulating growth and survival, how complete loss of these molecules in early BMMC progenitors (MCps) impacts maturation and growth during all phases of mast cell development is not fully understood. We show that the Lyn SFK and the p85α subunit of class I(A) PI3K play opposing roles in regulating the growth and maturation of BMMCs in part by regulating the level of PI3K. Loss of Lyn in BMMCs results in elevated PI3K activity and hyperactivation of AKT, which accelerates the rate of BMMC maturation due in part to impaired binding and phosphorylation of SHIP via Lyn's unique domain. In the absence of Lyn's unique domain, BMMCs behave in a manner similar to that of Lyn- or SHIP-deficient BMMCs. Importantly, loss of p85α in Lyn-deficient BMMCs not only represses the hyperproliferation associated with the loss of Lyn but also represses their accelerated maturation. The accelerated maturation of BMMCs due to loss of Lyn is associated with increased expression of microphthalmia-associated transcription factor (Mitf), which is repressed in MCps deficient in the expression of both Lyn and p85α relative to controls. Our results demonstrate a crucial interplay of Lyn, SHIP, and p85α in regulating the normal growth and maturation of BMMCs, in part by regulating the activation of AKT and the expression of Mitf.
Figures
References
-
- Anderson S. M., Jorgensen B. 1995. Activation of src-related tyrosine kinases by IL-3. J. Immunol. 155:1660–1670 - PubMed
-
- Brown M. T., Cooper J. A. 1996. Regulation, substrates and functions of src. Biochim. Biophys. Acta 1287:121–149 - PubMed
-
- Costa J. J., Weller P. F., Galli S. J. 1997. The cells of the allergic response: mast cells, basophils, and eosinophils. JAMA 278:1815–1822 - PubMed
-
- Damen J. E., Ware M. D., Kalesnikoff J., Hughes M. R., Krystal G. 2001. SHIP's C-terminus is essential for its hydrolysis of PI3P and inhibition of mast cell degranulation. Blood 97:1343–1351 - PubMed
-
- Durand B., et al. 1994. Long-term generation of human mast cells in serum-free cultures of CD34+ cord blood cells stimulated with stem cell factor and interleukin-3. Blood 84:3667–3674 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous