

Critical role of serpinB1 in regulating inflammatory responses in pulmonary influenza infection
- PMID: 21791661
- PMCID: PMC3144176
- DOI: 10.1093/infdis/jir352
Critical role of serpinB1 in regulating inflammatory responses in pulmonary influenza infection
Abstract
Background: Excessive inflammatory host response increases morbidity and mortality associated with seasonal respiratory influenza, and highly pathogenic virus strains are characterized by massive infiltration of monocytes and/or macrophages that produce a storm of injurious cytokines.
Methods: Here, we examined the role in respiratory influenza of serpinB1, an endogenous inhibitor of the serine proteases elastase, cathepsin G, and proteinase-3, increasingly recognized as regulators of inflammation.
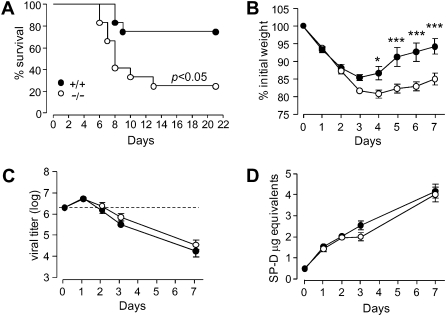
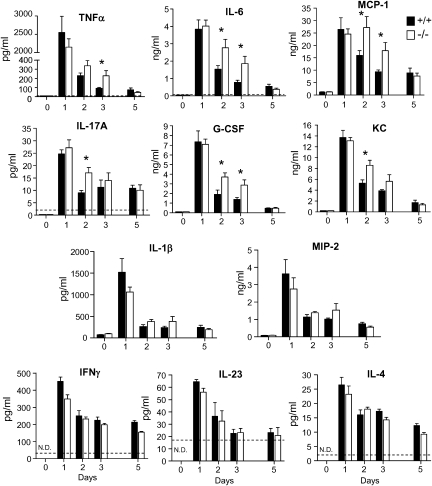
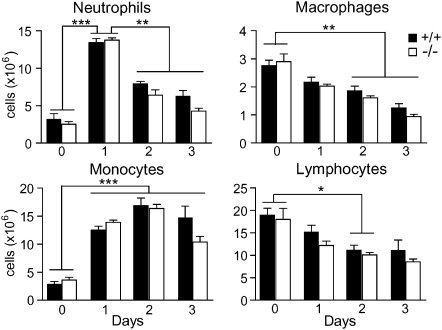
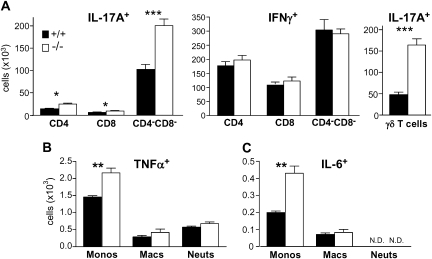
Results: After challenge with high-dose surfactant protein-D (SP-D)-sensitive influenza A/Philadelphia/82 (H3N2), serpinB1(-/-) mice died earlier and in greater numbers than did wild-type mice. Sublethally infected animals suffered increased morbidity, delayed resolution of epithelial injury, and increased immune cell death. Viral clearance and SP-D/SP-A upregulation were unimpaired and so were early virus-induced cytokine and chemokine burst and influx of large numbers of neutrophils and monocytes. Whereas initial cytokines and chemokines rapidly cleared in wild-type mice, TNF-α, IL-6, KC/CXCL1, G-CSF, IL-17A, and MCP-1/CCL2 remained elevated in serpinB1(-/-) mice. Monocyte-derived cells were the dominant immune cells in influenza-infected lungs, and those from serpinB1(-/-) mice produced excessive IL-6 and TNF-α when tested ex vivo. Pulmonary γδ T-cells that produced IL-17A were also increased.
Conclusions: Because viral clearance was unimpaired, the study highlights the critical role of serpinB1 in mitigating inflammation and restricting pro-inflammatory cytokine production in influenza infection.
Figures

References
-
- Pham CT. Neutrophil serine proteases: specific regulators of inflammation. Nat Rev Immunol. 2006;6:541–50. - PubMed
-
- Cooley J, Takayama TK, Shapiro SD, Schechter NM, Remold-O'Donnell E. The serpin MNEI inhibits elastase-like and chymotrypsin-like serine proteases through efficient reactions at two active sites. Biochemistry. 2001;40:15762–70. - PubMed
-
- Tecle T, White MR, Gantz D, Crouch EC, Hartshorn KL. Human neutrophil defensins increase neutrophil uptake of influenza A virus and bacteria and modify virus-induced respiratory burst responses. J Immunol. 2007;178:8046–52. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
