

Epstein-Barr virus latent membrane protein 1 (LMP1) C-terminal-activating region 3 contributes to LMP1-mediated cellular migration via its interaction with Ubc9
- PMID: 21795333
- PMCID: PMC3196420
- DOI: 10.1128/JVI.05035-11
Epstein-Barr virus latent membrane protein 1 (LMP1) C-terminal-activating region 3 contributes to LMP1-mediated cellular migration via its interaction with Ubc9
Abstract
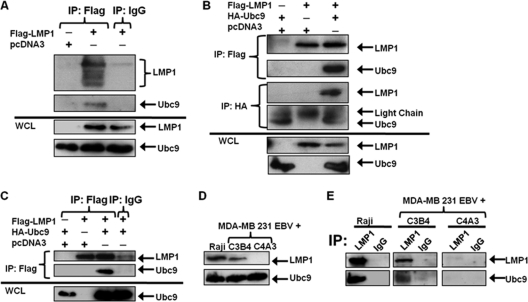
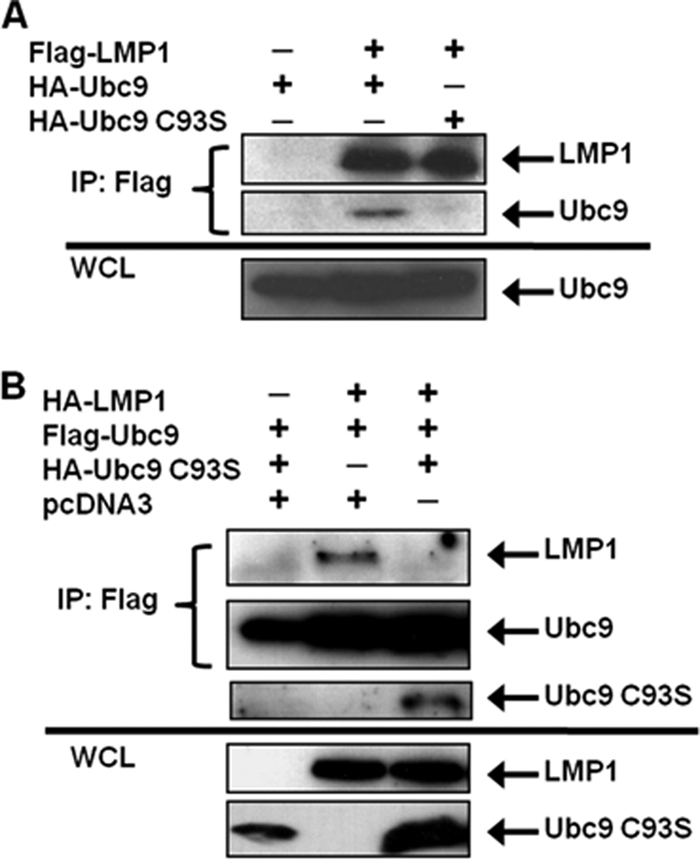
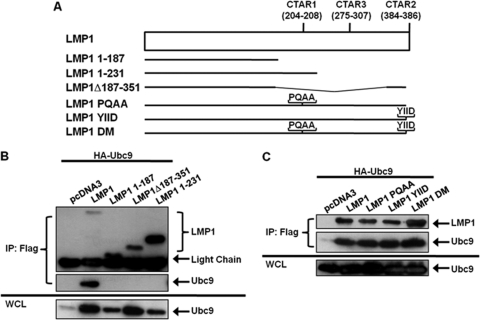
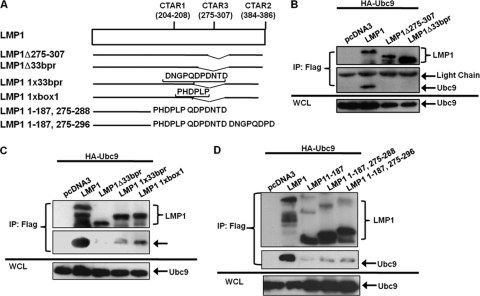
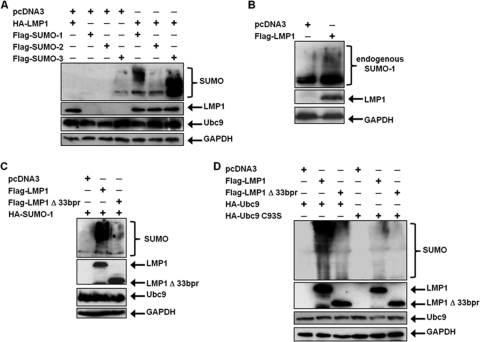
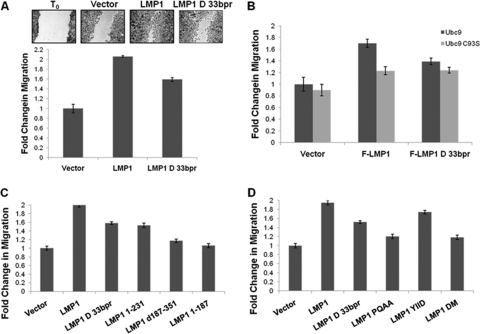
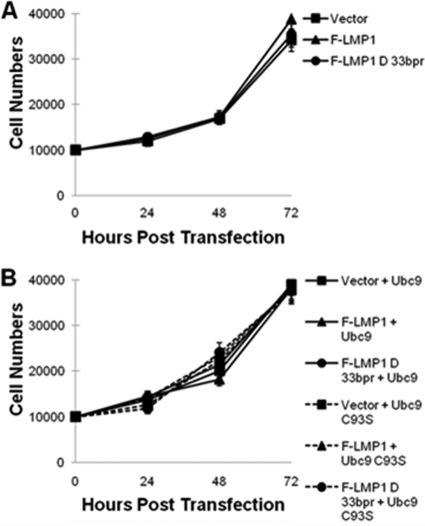
Epstein-Barr virus (EBV) latent membrane protein 1 (LMP1), the principal viral oncoprotein and a member of the tumor necrosis factor receptor superfamily, is a constitutively active membrane signaling protein that regulates multiple signal transduction pathways via its C-terminal-activating region 1 (CTAR1) and CTAR2, and also the less-studied CTAR3. Because protein sumoylation among other posttranslational modifications may regulate many signaling pathways induced by LMP1, we investigated whether during EBV latency LMP1 regulates sumoylation processes that control cellular activation and cellular responses. By immunoprecipitation experiments, we show that LMP1 interacts with Ubc9, the single reported SUMO-conjugating enzyme. Requirements for LMP1-Ubc9 interactions include enzymatically active Ubc9: expression of inactive Ubc9 (Ubc9 C93S) inhibited the LMP1-Ubc9 interaction. LMP1 CTAR3, but not CTAR1 and CTAR2, participated in the LMP1-Ubc9 interaction, and amino acid sequences found in CTAR3, including the JAK-interacting motif, contributed to this interaction. Furthermore, LMP1 expression coincided with increased sumoylation of cellular proteins, and disruption of the Ubc9-LMP1 CTAR3 interaction almost completely abrogated LMP1-induced protein sumoylation, suggesting that this interaction promotes the sumoylation of downstream targets. Additional consequences of the disruption of the LMP1 CTAR3-Ubc9 interaction revealed effects on cellular migration, a hallmark of oncogenesis. Together, these data demonstrate that LMP1 CTAR3 does in fact function in intracellular signaling and leads to biological effects. We propose that LMP1, by interaction with Ubc9, modulates sumoylation processes, which regulate signal transduction pathways that affect phenotypic changes associated with oncogenesis.
Figures
References
-
- Albuquerque M. L., Waters C. M., Savla U., Schnaper H. W., Flozak A. S. 2000. Shear stress enhances human endothelial cell wound closure in vitro. Am. J. Physiol. Heart Circ. Physiol. 279:H293–H302 - PubMed
-
- Aviel S., Winberg G., Massucci M., Ciechanover A. 2000. Degradation of the Epstein-Barr virus latent membrane protein 1 (LMP1) by the ubiquitin-proteasome pathway. Targeting via ubiquitination of the N-terminal residue. J. Biol. Chem. 275:23491–23499 - PubMed
-
- Brennan P., Floettmann J. E., Mehl A., Jones M., Rowe M. 2001. Mechanism of action of a novel latent membrane protein-1 dominant negative. J. Biol. Chem. 276:1195–1203 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
