

The budding yeast polo-like kinase Cdc5 regulates the Ndt80 branch of the meiotic recombination checkpoint pathway
- PMID: 21795394
- PMCID: PMC3172271
- DOI: 10.1091/mbc.E11-06-0482
The budding yeast polo-like kinase Cdc5 regulates the Ndt80 branch of the meiotic recombination checkpoint pathway
Abstract
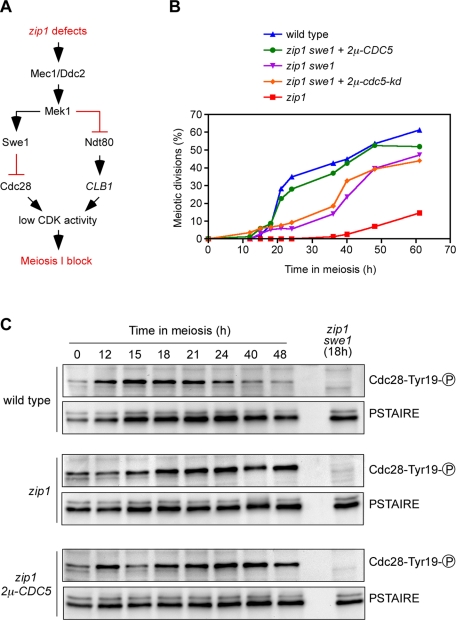
Defects in chromosome synapsis and/or meiotic recombination activate a surveillance mechanism that blocks meiotic cell cycle progression to prevent anomalous chromosome segregation and formation of aberrant gametes. In the budding yeast zip1 mutant, which lacks a synaptonemal complex component, the meiotic recombination checkpoint is triggered, resulting in extremely delayed meiotic progression. We report that overproduction of the polo-like kinase Cdc5 partially alleviates the meiotic prophase arrest of zip1, leading to the formation of inviable meiotic products. Unlike vegetative cells, we demonstrate that Cdc5 overproduction does not stimulate meiotic checkpoint adaptation because the Mek1 kinase remains activated in zip1 2μ-CDC5 cells. Inappropriate meiotic divisions in zip1 promoted by high levels of active Cdc5 do not result from altered function of the cyclin-dependent kinase (CDK) inhibitor Swe1. In contrast, CDC5 overexpression leads to premature induction of the Ndt80 transcription factor, which drives the expression of genes required for meiotic divisions, including CLB1. We also show that depletion of Cdc5 during meiotic prophase prevents the production of Ndt80 and that CDK activity contributes to the induction of Ndt80 in zip1 cells overexpressing CDC5. Our results reveal a role for Cdc5 in meiotic checkpoint control by regulating Ndt80 function.
Figures







References
-
- Amon A, Surana U, Muroff I, Nasmyth K. Regulation of p34CDC28 tyrosine phosphorylation is not required for entry into mitosis in S. cerevisiae. Nature. 1992;355:368–371. - PubMed
-
- Archambault V, Glover DM. Polo-like kinases: conservation and divergence in their functions and regulation. Nat Rev Mol Cell Biol. 2009;10:265–275. - PubMed
-
- Bailis JM, Roeder GS. Pachytene exit controlled by reversal of Mek1-dependent phosphorylation. Cell. 2000;101:211–221. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
