

Optogenetic interrogation of dopaminergic modulation of the multiple phases of reward-seeking behavior
- PMID: 21795535
- PMCID: PMC3171183
- DOI: 10.1523/JNEUROSCI.2246-11.2011
Optogenetic interrogation of dopaminergic modulation of the multiple phases of reward-seeking behavior
Abstract
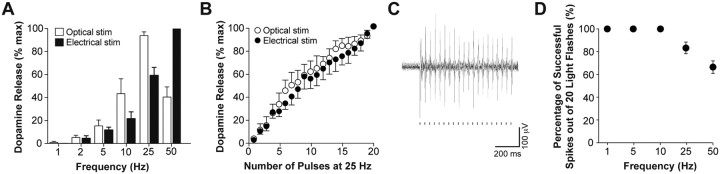
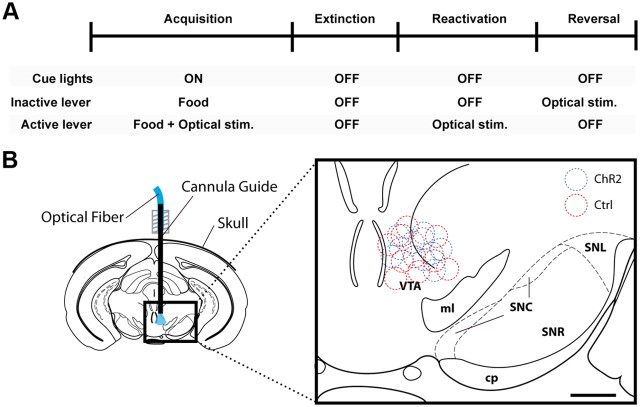
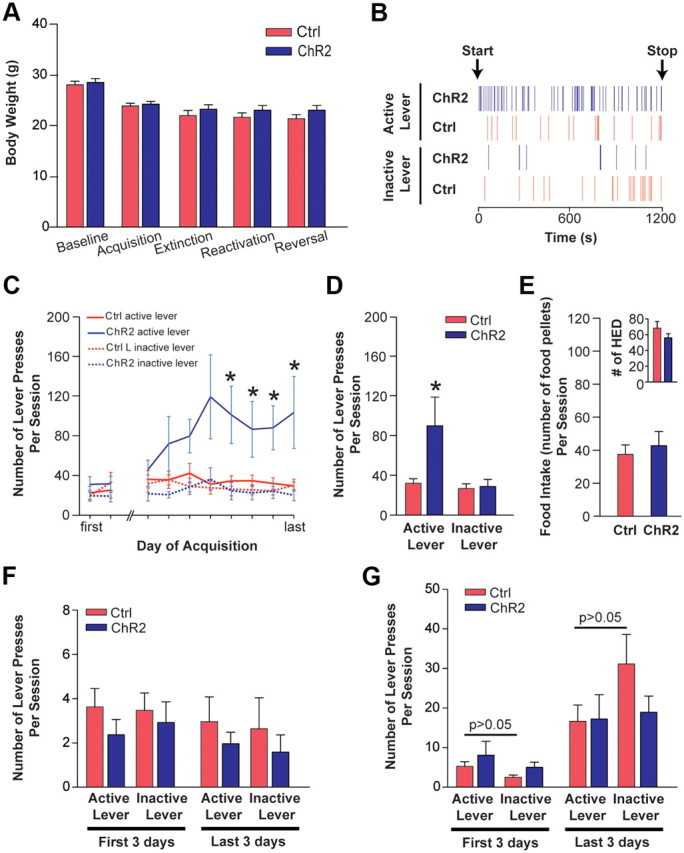
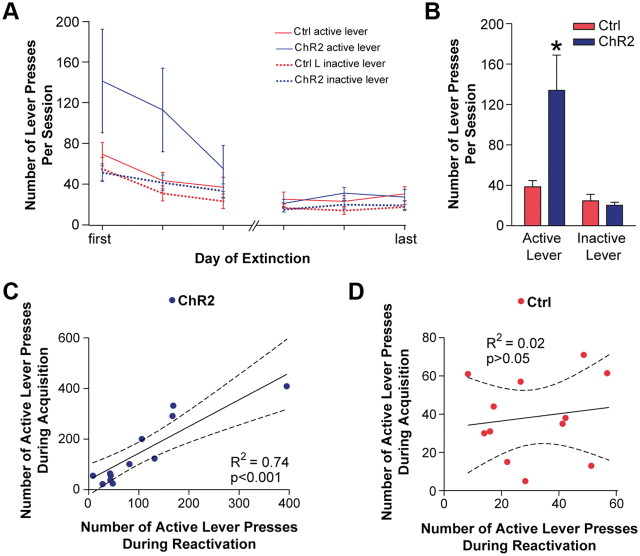
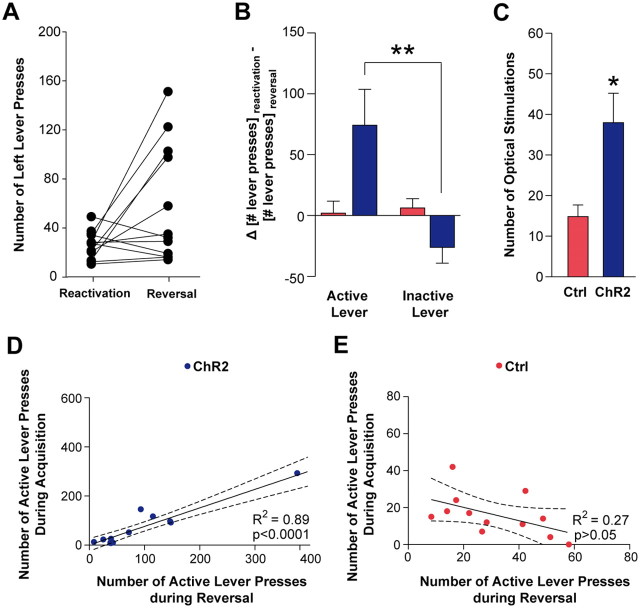
Phasic activation of dopaminergic neurons is associated with reward-predicting cues and supports learning during behavioral adaptation. While noncontingent activation of dopaminergic neurons in the ventral tegmental are (VTA) is sufficient for passive behavioral conditioning, it remains unknown whether the phasic dopaminergic signal is truly reinforcing. In this study, we first targeted the expression of channelrhodopsin-2 to dopaminergic neurons of the VTA and optimized optogenetically evoked dopamine transients. Second, we showed that phasic activation of dopaminergic neurons in freely moving mice causally enhances positive reinforcing actions in a food-seeking operant task. Interestingly, such effect was not found in the absence of food reward. We further found that phasic activation of dopaminergic neurons is sufficient to reactivate previously extinguished food-seeking behavior in the absence of external cues. This was also confirmed using a single-session reversal paradigm. Collectively, these data suggest that activation of dopaminergic neurons facilitates the development of positive reinforcement during reward-seeking and behavioral flexibility.
Figures
Comment in
-
Shedding light on the role of ventral tegmental area dopamine in reward.J Neurosci. 2011 Dec 14;31(50):18195-7. doi: 10.1523/JNEUROSCI.4924-11.2011. J Neurosci. 2011. PMID: 22171025 Free PMC article. No abstract available.
References
-
- Berridge KC. The debate over dopamine's role in reward: the case for incentive salience. Psychopharmacology (Berl) 2007;191:391–431. - PubMed
-
- Blaha CD, Phillips AG. Application of in vivo electrochemistry to the measurement of changes in dopamine release during intracranial self-stimulation. J Neurosci Methods. 1990;34:125–133. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases