

Synaptic and morphological neuroadaptations in the putamen associated with long-term, relapsing alcohol drinking in primates
- PMID: 21796110
- PMCID: PMC3194078
- DOI: 10.1038/npp.2011.140
Synaptic and morphological neuroadaptations in the putamen associated with long-term, relapsing alcohol drinking in primates
Abstract
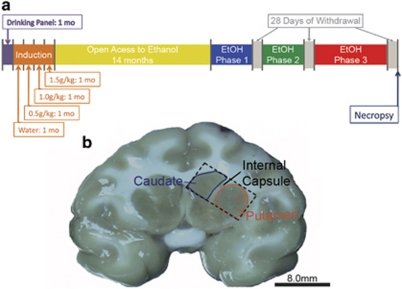
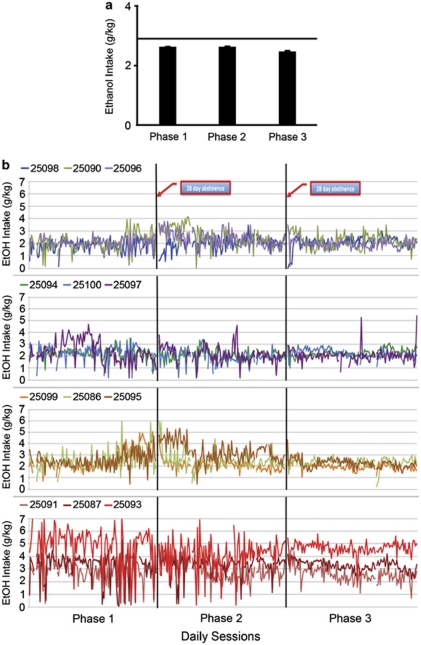
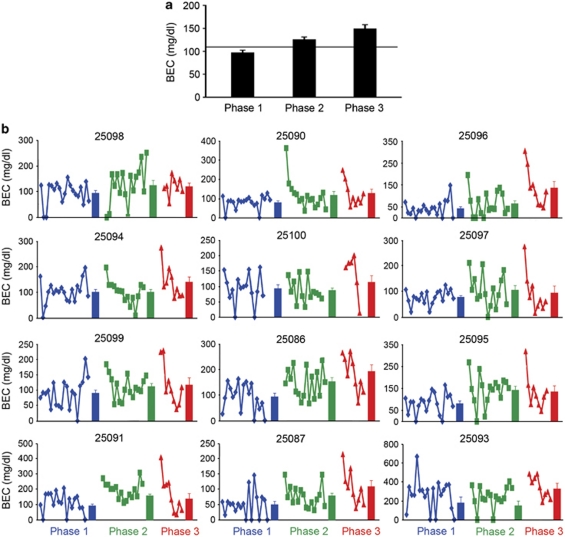
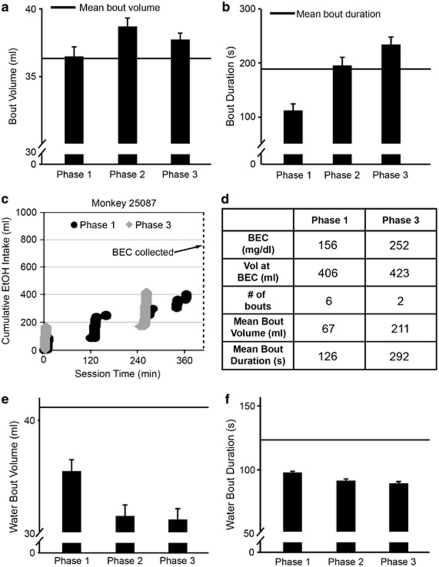
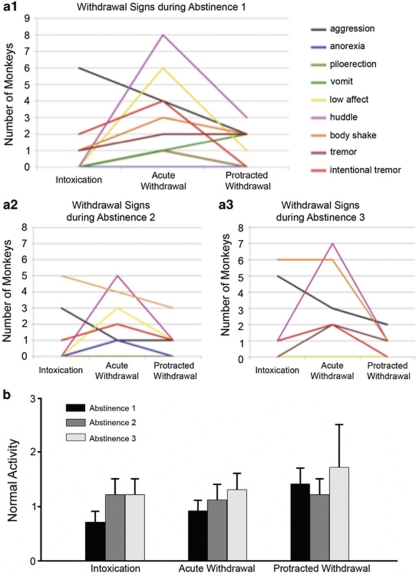
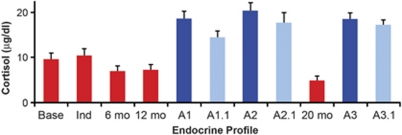
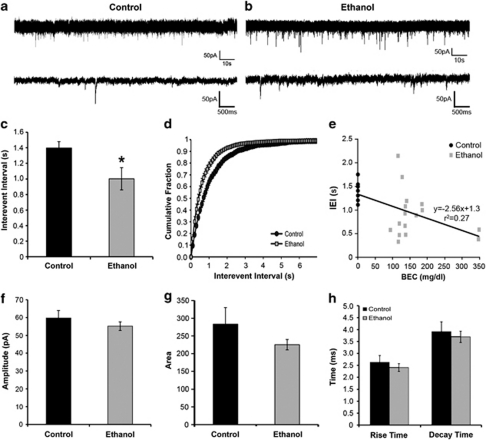
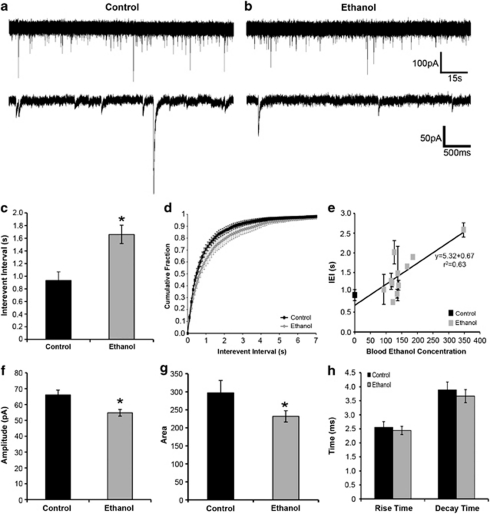
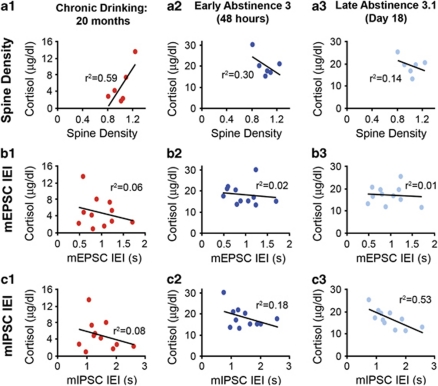
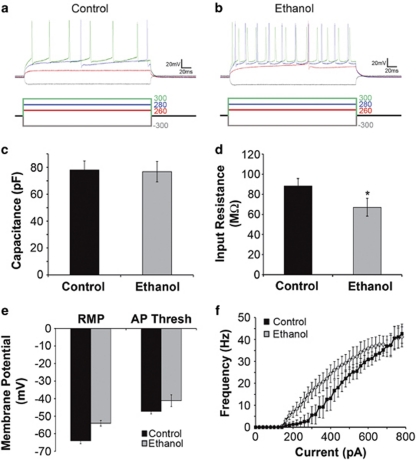
Alcoholism and alcohol use disorders are characterized by several months to decades of heavy and problematic drinking, interspersed with periods of abstinence and relapse to heavy drinking. This alcohol-drinking phenotype was modeled using macaque monkeys to explore neuronal adaptations in the striatum, a brain region controlling habitual behaviors. Prolonged drinking with repeated abstinence narrowed the variability in daily intake, increased the amount of ethanol consumed in bouts, and led to higher blood ethanol concentrations more than twice the legal intoxication limit. After the final abstinence period of this extensive drinking protocol, we found a selective increase in dendritic spine density and enhanced glutamatergic transmission in the putamen, but not in the caudate nucleus. Intrinsic excitability of medium-sized spiny neurons was also enhanced in the putamen of alcohol-drinking monkeys in comparison with non-drinkers, and GABAeric transmission was selectively suppressed in the putamen of heavy drinkers. These morphological and physiological changes indicate a shift in the balance of inhibitory/excitatory transmission that biases the circuit toward an enduring increase in synaptic activation of putamen output as a consequence of prolonged heavy drinking/relapse. The resultant potential for increased putamen activation may underlie an alcohol-drinking phenotype of regulated drinking and sustained intoxication.
Figures

References
-
- Becker HC, Lopez MF. Increased ethanol drinking after repeated chronic ethanol exposure and withdrawal experience in C57BL/6J mice. Alcohol Clin Exp Res. 2004;28:1829–1838. - PubMed
-
- Carden WB, Alexander GM, Friedman DP, Daunais JB, Grant KA, Mu J, et al. Chronic ethanol drinking reduces native T-type calcium current in the thalamus of nonhuman primates. Brain Res. 2006;1089:92–100. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
