

Impaired respiratory and body temperature control upon acute serotonergic neuron inhibition
- PMID: 21798952
- PMCID: PMC3729433
- DOI: 10.1126/science.1205295
Impaired respiratory and body temperature control upon acute serotonergic neuron inhibition
Abstract
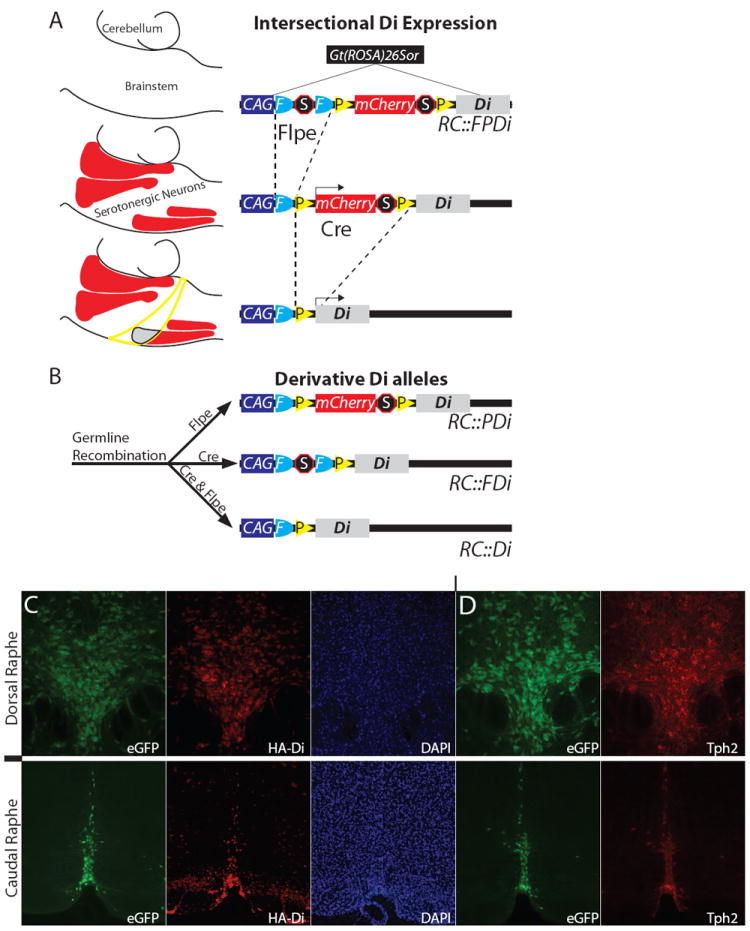
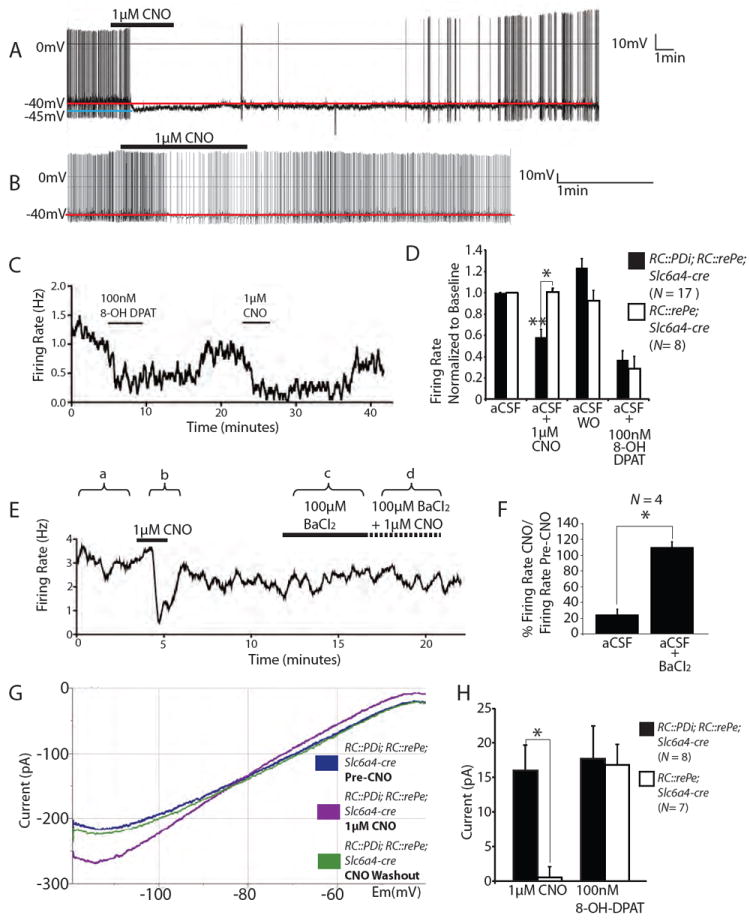
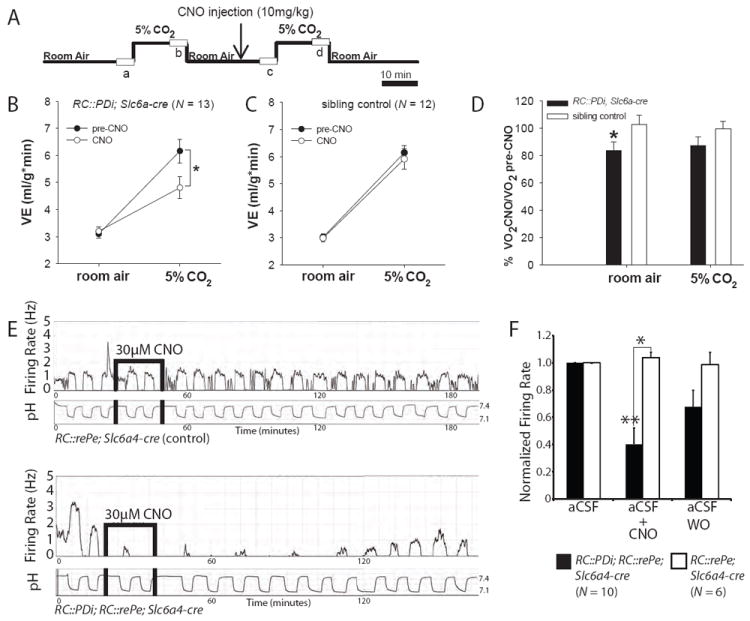
Physiological homeostasis is essential for organism survival. Highly responsive neuronal networks are involved, but their constituent neurons are just beginning to be resolved. To query brain serotonergic neurons in homeostasis, we used a neuronal silencing tool, mouse RC::FPDi (based on the synthetic G protein-coupled receptor Di), designed for cell type-specific, ligand-inducible, and reversible suppression of action potential firing. In mice harboring Di-expressing serotonergic neurons, administration of the ligand clozapine-N-oxide (CNO) by systemic injection attenuated the chemoreflex that normally increases respiration in response to tissue carbon dioxide (CO(2)) elevation and acidosis. At the cellular level, CNO suppressed firing rate increases evoked by CO(2) acidosis. Body thermoregulation at room temperature was also disrupted after CNO triggering of Di; core temperatures plummeted, then recovered. This work establishes that serotonergic neurons regulate life-sustaining respiratory and thermoregulatory networks, and demonstrates a noninvasive tool for mapping neuron function.
Conflict of interest statement
The authors declare no conflict of interests
Figures

Comment in
-
Comment on "Impaired respiratory and body temperature control upon acute serotonergic neuron inhibition".Science. 2012 Aug 10;337(6095):646; author reply 646. doi: 10.1126/science.1222519. Science. 2012. PMID: 22879486
References
-
- Leusen IR. Chemosensitivity of the respiratory center; influence of CO2 in the cerebral ventricles on respiration. Am J Physiol. 1954 Jan;176:39. - PubMed
-
- Kety SS, Forster RE, Comroe Julius H., Jr March 13, 1911-July 31, 1984. Biogr Mem Natl Acad Sci. 2001;79:66. - PubMed
-
- Smith CA, Rodman JR, Chenuel BJ, Henderson KS, Dempsey JA. Response time and sensitivity of the ventilatory response to CO2 in unanesthetized intact dogs: central vs. peripheral chemoreceptors. J Appl Physiol. 2006 Jan;100:13. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 5R21MH083613-02/MH/NIMH NIH HHS/United States
- 5R21DA023643-02/DA/NIDA NIH HHS/United States
- 5R01HL028066-30/HL/NHLBI NIH HHS/United States
- 5R01HD052772/HD/NICHD NIH HHS/United States
- R01 HD052772/HD/NICHD NIH HHS/United States
- R21 MH083613/MH/NIMH NIH HHS/United States
- 5P01HD036379-13/HD/NICHD NIH HHS/United States
- F31 NS073276/NS/NINDS NIH HHS/United States
- R21 DA023643/DA/NIDA NIH HHS/United States
- F32 HD063257/HD/NICHD NIH HHS/United States
- P01 HD036379/HD/NICHD NIH HHS/United States
- R01 HL028066/HL/NHLBI NIH HHS/United States
- F32HD063257-01A1/HD/NICHD NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
