

PGC-1β deficiency accelerates the transition to heart failure in pressure overload hypertrophy
- PMID: 21799152
- PMCID: PMC3175248
- DOI: 10.1161/CIRCRESAHA.111.243964
PGC-1β deficiency accelerates the transition to heart failure in pressure overload hypertrophy
Abstract
Rationale: Pressure overload cardiac hypertrophy, a risk factor for heart failure, is associated with reduced mitochondrial fatty acid oxidation (FAO) and oxidative phosphorylation (OXPHOS) proteins that correlate in rodents with reduced PGC-1α expression.
Objective: To determine the role of PGC-1β in maintaining mitochondrial energy metabolism and contractile function in pressure overload hypertrophy.
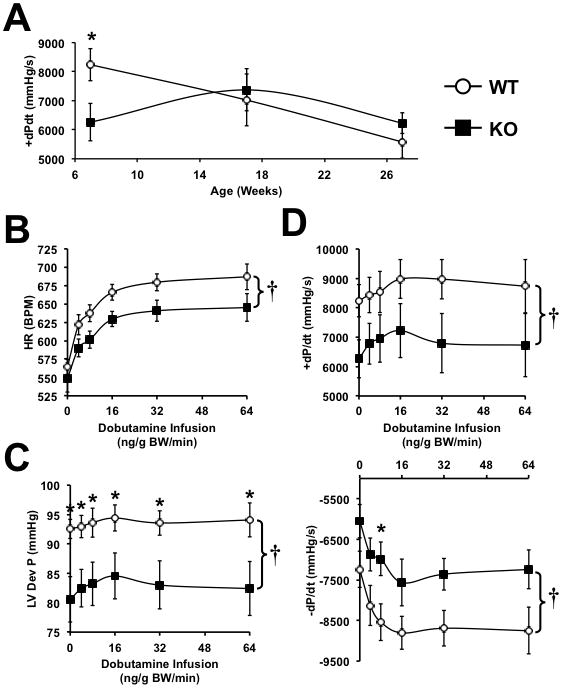
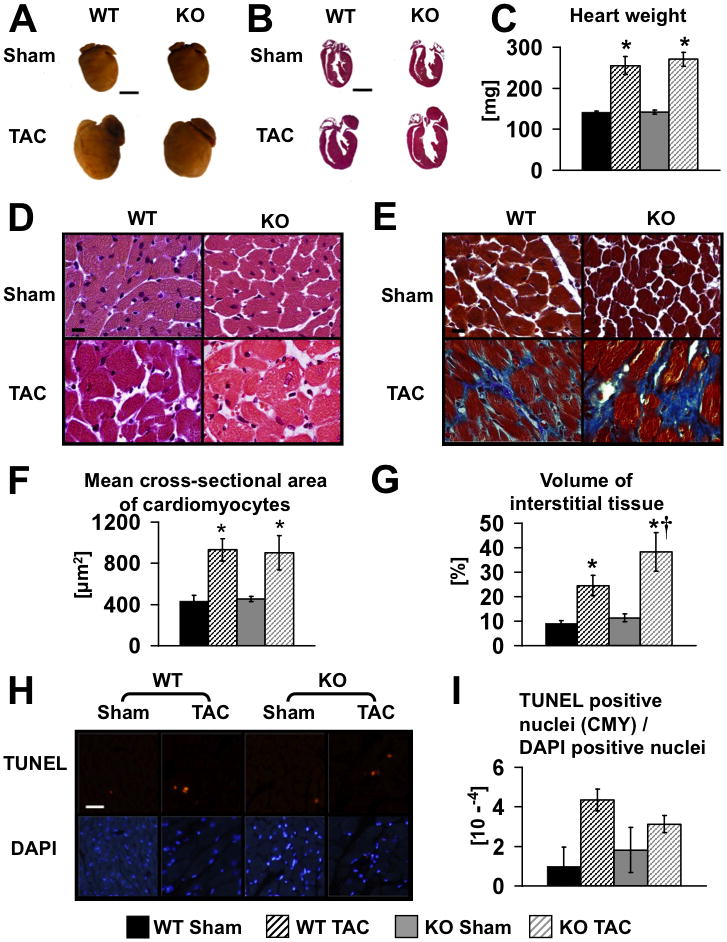
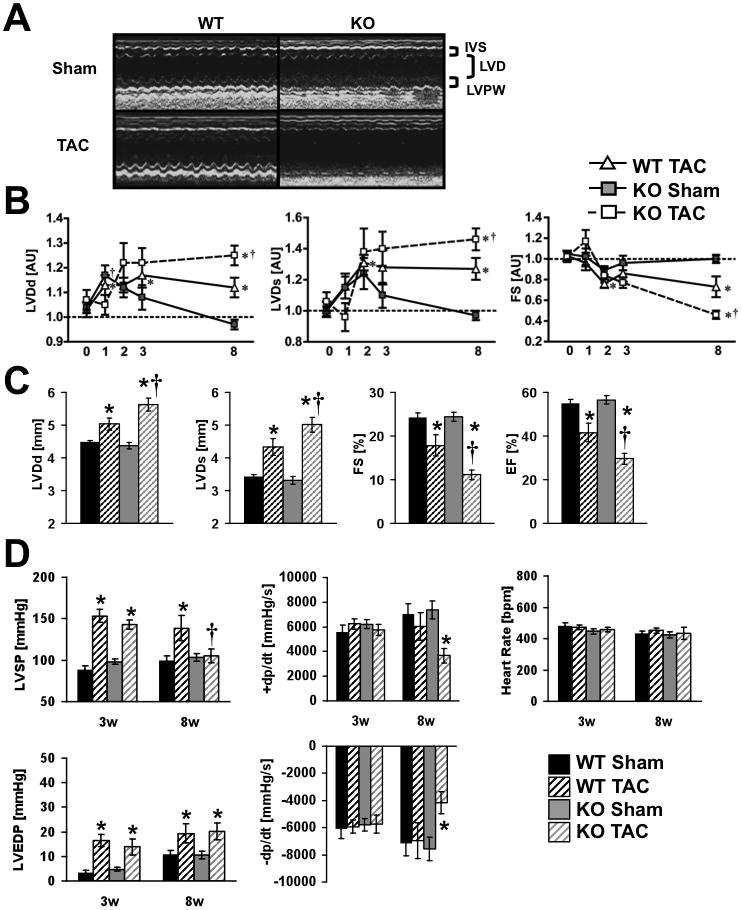
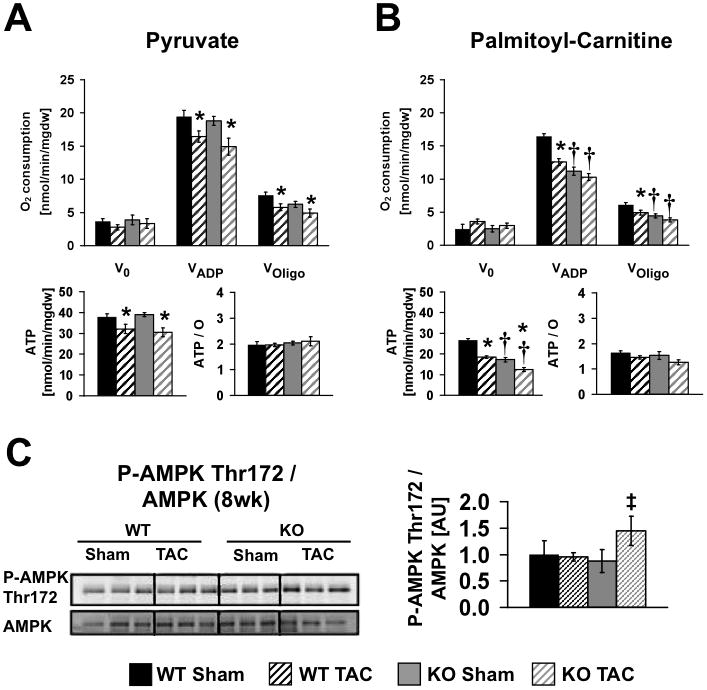
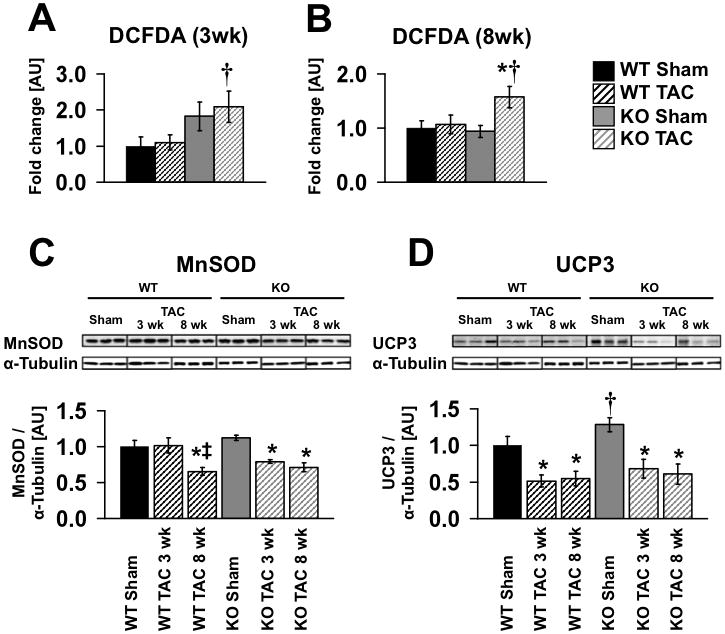
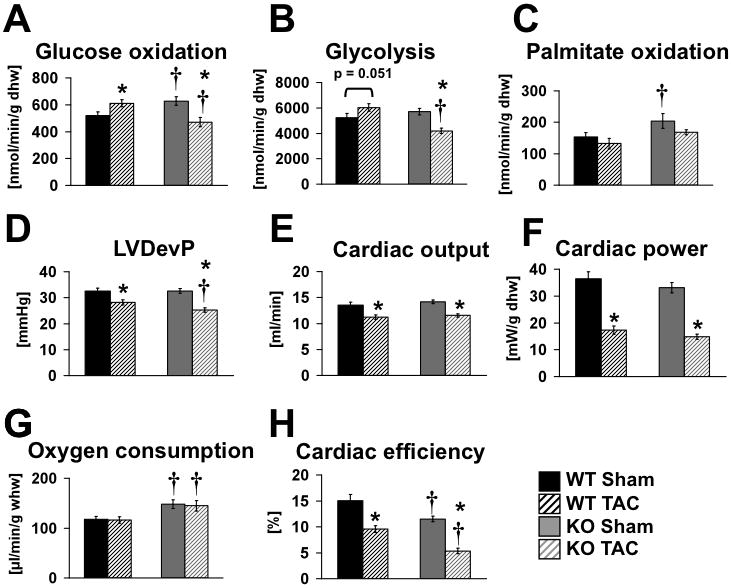
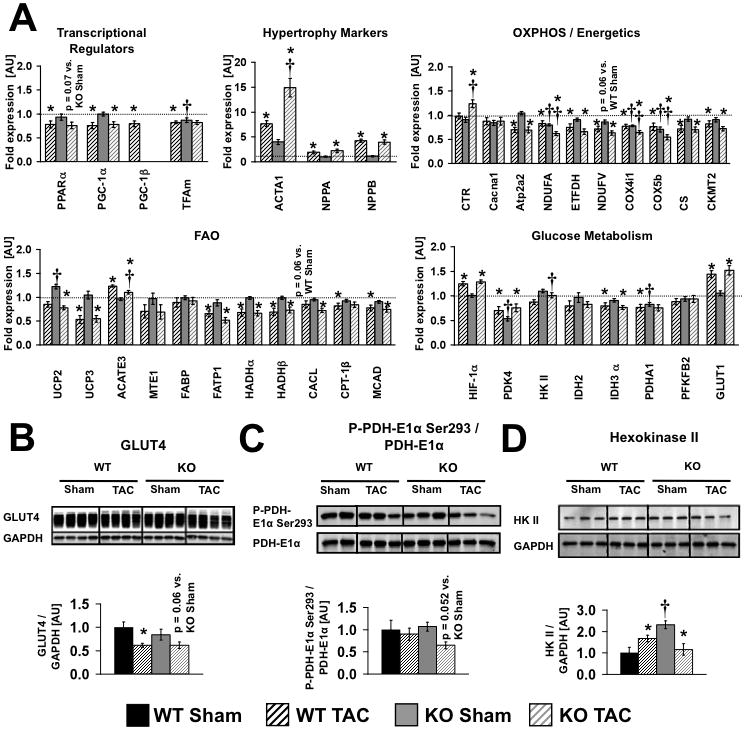
Methods and results: PGC-1β deficient (KO) mice and wildtype (WT) controls were subjected to transverse aortic constriction (TAC). Although LV function was modestly reduced in young KO hearts, there was no further decline with age so that LV function was similar between KO and WT when TAC was performed. WT-TAC mice developed relatively compensated LVH, despite reduced mitochondrial function and repression of OXPHOS and FAO genes. In nonstressed KO hearts, OXPHOS gene expression and palmitoyl-carnitine-supported mitochondrial function were reduced to the same extent as banded WT, but FAO gene expression was normal. Following TAC, KO mice progressed more rapidly to heart failure and developed more severe mitochondrial dysfunction, despite a similar overall pattern of repression of OXPHOS and FAO genes as WT-TAC. However, in relation to WT-TAC, PGC-1β deficient mice exhibited greater degrees of oxidative stress, decreased cardiac efficiency, lower rates of glucose metabolism, and repression of hexokinase II protein.
Conclusions: PGC-1β plays an important role in maintaining baseline mitochondrial function and cardiac contractile function following pressure overload hypertrophy by preserving glucose metabolism and preventing oxidative stress.
Conflict of interest statement
C. Lelliott is employed by Astra Zeneca. The other authors have no conflicts of interest to disclose.
Figures
References
-
- Meirhaeghe A, Crowley V, Lenaghan C, Lelliott C, Green K, Stewart A, Hart K, Schinner S, Sethi JK, Yeo G, Brand MD, Cortright RN, O’Rahilly S, Montague C, Vidal-Puig AJ. Characterization of the human, mouse and rat pgc1 beta (peroxisome-proliferator-activated receptor-gamma co-activator 1 beta) gene in vitro and in vivo. Biochem J. 2003;373:155–165. - PMC - PubMed
-
- Lin J, Tarr PT, Yang R, Rhee J, Puigserver P, Newgard CB, Spiegelman BM. Pgc-1beta in the regulation of hepatic glucose and energy metabolism. J Biol Chem. 2003;278:30843–30848. - PubMed
-
- Lehman JJ, Boudina S, Banke NH, Sambandam N, Han X, Young DM, Leone TC, Gross RW, Lewandowski ED, Abel ED, Kelly DP. The transcriptional coactivator pgc-1alpha is essential for maximal and efficient cardiac mitochondrial fatty acid oxidation and lipid homeostasis. Am J Physiol Heart Circ Physiol. 2008;295:H185–196. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
