

Coenzyme Q10 decreases amyloid pathology and improves behavior in a transgenic mouse model of Alzheimer's disease
- PMID: 21799249
- PMCID: PMC3267988
- DOI: 10.3233/JAD-2011-110209
Coenzyme Q10 decreases amyloid pathology and improves behavior in a transgenic mouse model of Alzheimer's disease
Abstract
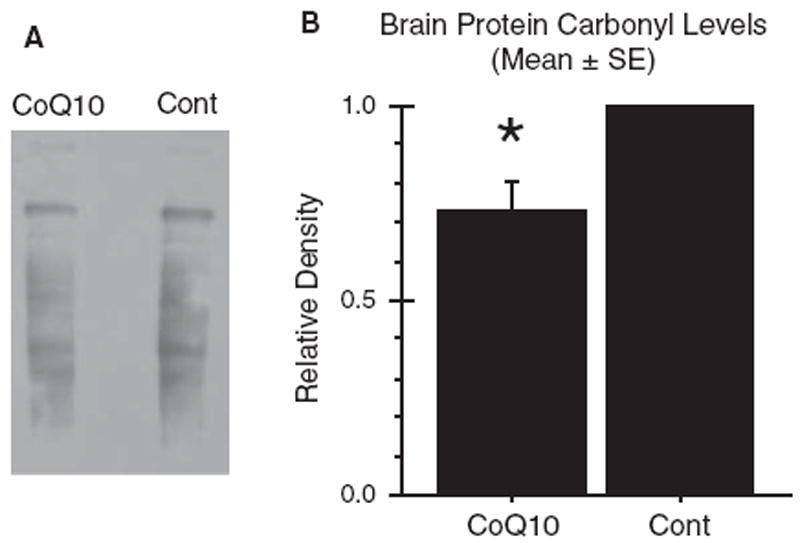
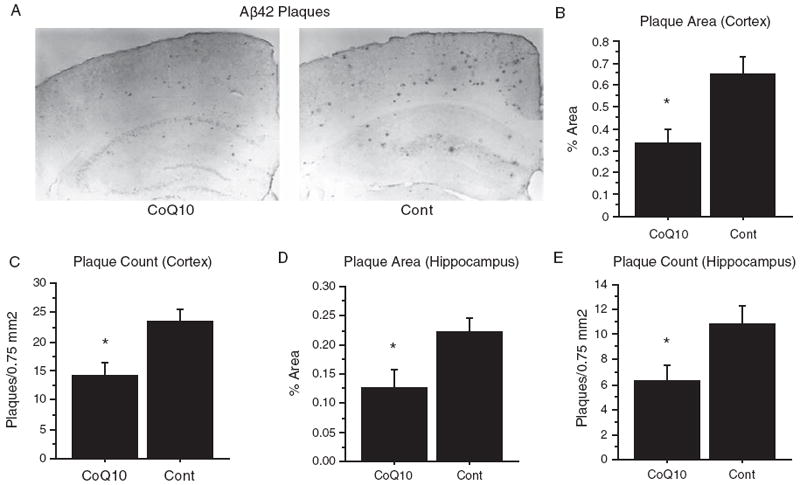
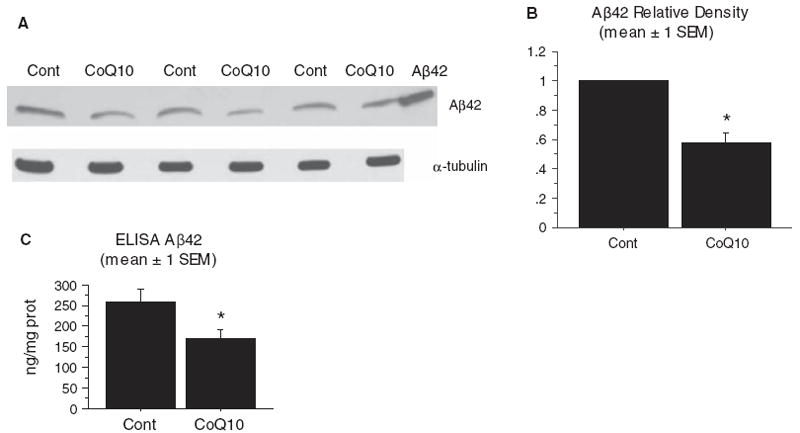
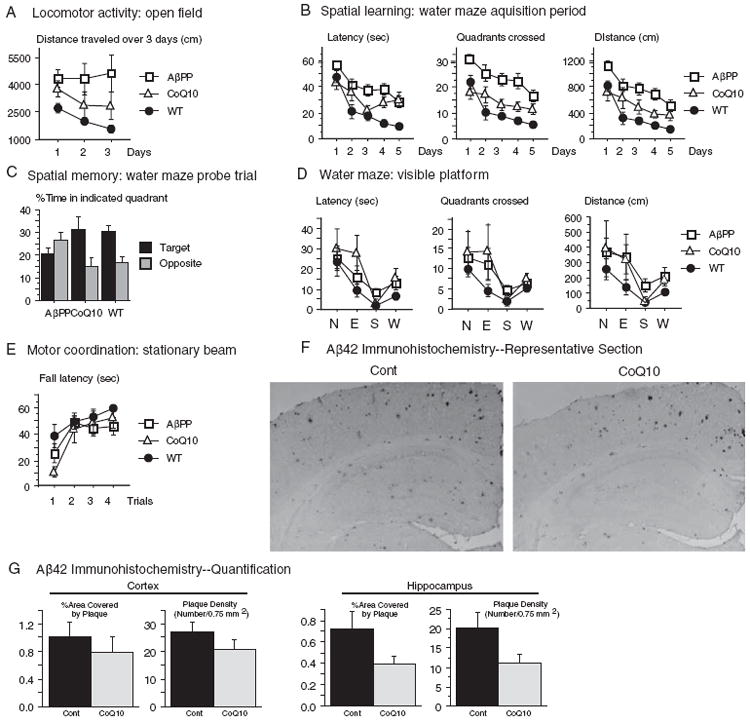
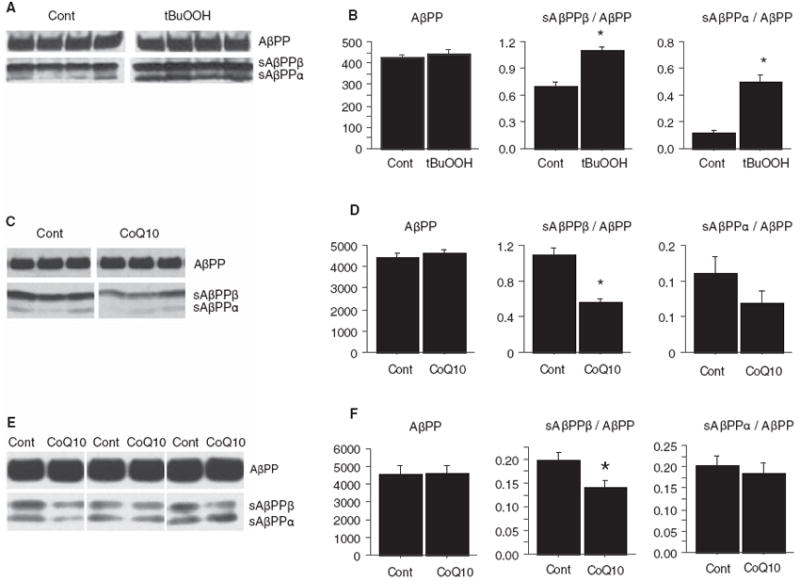
Increased oxidative stress is implicated in the pathogenesis of Alzheimer's disease (AD). A large body of evidence suggests that mitochondrial dysfunction and increased reactive oxygen species occur prior to amyloid-β (Aβ) deposition. Coenzyme Q10 (CoQ10), a component of the mitochondrial electron transport chain, is well characterized as a neuroprotective antioxidant in animal models and human trials of Huntington's disease and Parkinson's disease, and reduces plaque burden in AβPP/PS1 mice. We now show that CoQ10 reduces oxidative stress and amyloid pathology and improves behavioral performance in the Tg19959 mouse model of AD. CoQ10 treatment decreased brain levels of protein carbonyls, a marker of oxidative stress. CoQ10 treatment resulted in decreased plaque area and number in hippocampus and in overlying cortex immunostained with an Aβ42-specific antibody. Brain Aβ42 levels were also decreased by CoQ10 supplementation. Levels of amyloid-β protein precursor (AβPP) β-carboxyterminal fragments were decreased. Importantly, CoQ10-treated mice showed improved cognitive performance during Morris water maze testing. Our results show decreased pathology and improved behavior in transgenic AD mice treated with the naturally occurring antioxidant compound CoQ10. CoQ10 is well tolerated in humans and may be promising for therapeutic trials in AD.
Figures
References
-
- Drake J, Link CD, Butterfield DA. Oxidative stress precedes fibrillar deposition of Alzheimer’s disease amyloid beta-peptide (1-42) in a transgenic Caenorhabditis elegans model. Neurobiol Aging. 2003;24:415–420. - PubMed
-
- Keller JN, Schmitt FA, Scheff SW, Ding Q, Chen Q, Butterfield DA, Markesbery WR. Evidence of increased oxidative damage in subjects with mild cognitive impairment. Neurology. 2005;64:1152–1156. - PubMed
-
- Mecocci P, MacGarvey U, Beal MF. Oxidative damage to mitochondrial DNA is increased in Alzheimer’s disease. Ann Neurol. 1994;36:747–751. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
