

Krüppel-like factor 6 expression changes during trophoblast syncytialization and transactivates ßhCG and PSG placental genes
- PMID: 21799854
- PMCID: PMC3142166
- DOI: 10.1371/journal.pone.0022438
Krüppel-like factor 6 expression changes during trophoblast syncytialization and transactivates ßhCG and PSG placental genes
Abstract
Background: Krüppel-like factor-6 (KLF6) is a widely expressed member of the Sp1/KLF family of transcriptional regulators involved in differentiation, cell cycle control and proliferation in several cell systems. Even though the highest expression level of KLF6 has been detected in human and mice placenta, its function in trophoblast physiology is still unknown.
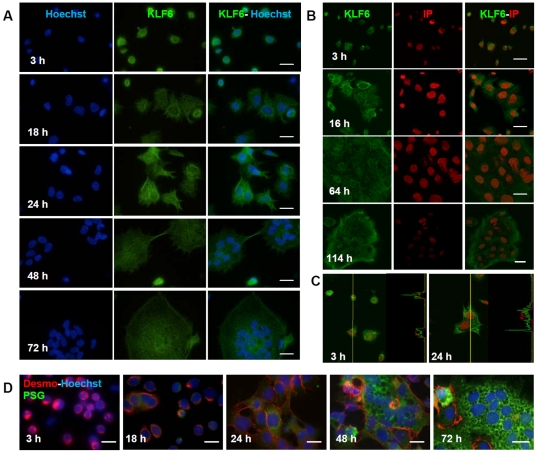
Methodology/principal findings: Herein, we explored KLF6 expression and sub-cellular distribution in human trophoblast cells differentiating into the syncytial pathway, and its role in the regulation of genes associated with placental development and pregnancy maintenance. Confocal immunofluorescence microscopy demonstrated that KLF6 is expressed throughout human cytotrophoblast differentiation showing no evident modifications in its nuclear and cytoplasmic localization pattern. KLF6 transcript and protein peaked early during the syncytialization process as determined by qRT-PCR and western blot assays. Overexpression of KLF6 in trophoblast-derived JEG-3 cells showed a preferential nuclear signal correlating with enhanced expression of human β-chorionic gonadotropin (βhCG) and pregnancy-specific glycoprotein (PSG) genes. Moreover, KLF6 transactivated βhCG5, PSG5 and PSG3 gene promoters. Deletion of KLF6 Zn-finger DNA binding domain or mutation of the consensus KLF6 binding site abolished transactivation of the PSG5 promoter.
Conclusions/significance: Results are consistent with KLF6 playing a role as transcriptional regulator of relevant genes for placental differentiation and physiology such as βhCG and PSG, in agreement with an early and transient increase of KLF6 expression during trophoblast syncytialization.
Conflict of interest statement
Figures



References
-
- Huppertz B, Borges M. Placenta trophoblast fusion. Methods Mol Biol. 2008;475:135–147. - PubMed
-
- Baczyk D, Drewlo S, Proctor L, Dunk C, Lye S, et al. Glial cell missing-1 transcription factor is required for the differentiation of the human trophoblast. Cell Death Differ. 2009;16:719–727. - PubMed
-
- Yang M, Lei ZM, Rao Ch V. The central role of human chorionic gonadotropin in the formation of human placental syncytium. Endocrinology. 2003;144:1108–1120. - PubMed
-
- Knofler M, Saleh L, Bauer S, Galos B, Rotheneder H, et al. Transcriptional regulation of the human chorionic gonadotropin beta gene during villous trophoblast differentiation. Endocrinology. 2004;145:1685–1694. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
